Triceratops
Triceratops horridus
"Horrible three-horned face"
About this species
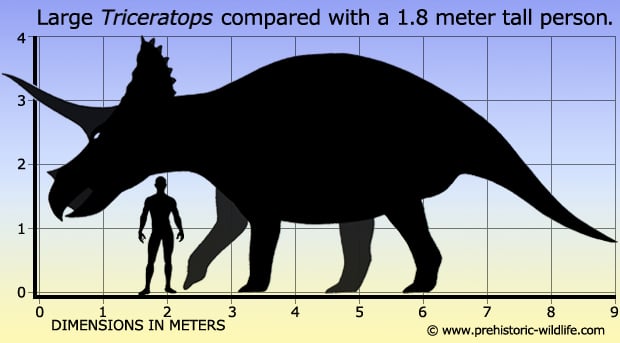
Triceratops horridus is one of the most recognizable dinosaurs of the Late Cretaceous, famous for its three facial horns and large bony neck frill. It lived between 68 and 66 million years ago in the Lance and Hell Creek Formations of the western United States. Up to 9 meters long and weighing up to 12 tonnes, it was one of the largest herbivores in its ecosystem. The skull could exceed 2.5 meters in length, ranking among the largest of any terrestrial animal. Its dental battery included 36 to 40 tooth columns per jaw side, enabling processing of large volumes of fibrous plant material. The horns and frill were likely used primarily for species recognition, sexual display, and dominance among individuals, with possible secondary use in defense against Tyrannosaurus rex.
Geological formation & environment
The Hell Creek Formation is an Upper Cretaceous sedimentary unit (Maastrichtian, 68 to 66 Ma) outcropping in Montana, North Dakota, South Dakota, and Wyoming. Deposited along the margins of the retreating Western Interior Seaway, the formation preserves one of the world's most complete dinosaur faunas, including Tyrannosaurus rex, Triceratops, Edmontosaurus, and Ankylosaurus. The Lance Formation is the chronological equivalent of Hell Creek in southern Wyoming and northern Colorado. Both formations end at the Cretaceous-Paleogene (K-Pg) boundary, marked by the mass extinction 66 Ma ago associated with the Chicxulub asteroid impact.
Image gallery















Life reconstruction of Triceratops horridus based on specimen P 256878, by Connor Ashbridge (2023).
Connor Ashbridge (Ddinodan), CC BY-SA 4.0
Ecology and behavior
Habitat
Triceratops lived on the coastal fluvial plains and interior forests of Laramidia, the western subcontinent of North America during the Maastrichtian. The Hell Creek Formation environment was dominated by meandering river floodplains, with dense cover of angiosperms, conifers, ferns, and palms. The climate was warm and seasonal, with moderate annual precipitation. Triceratops coexisted with Tyrannosaurus rex, Edmontosaurus, Ankylosaurus, and Torosaurus in this high-diversity faunal ecosystem (Campione & Evans, 2011). It was the dominant ceratopsid herbivore at the end of the Cretaceous, with wide distribution in the Hell Creek and Lance Formations. Population density estimates, based on fóssil abundance in systematic excavations, suggest it was one of the most numerous herbivores in the ecosystem, likely forming dispersed groups or herds.
Feeding
Triceratops was a herbivore with dentition specialized for cutting and grinding resistant vegetation. Its dental battery of 36 to 40 columns per jaw side consisted of continuously replacing teeth, a highly efficient adaptation for silica-rich or highly fibrous plants, such as palms, cycads, and rigid-stemmed angiosperms (Horner & Goodwin, 2006). The horny beak was robust and designed for cutting plant stems. Stable isotope analyses of teeth suggest a generalist C3 plant diet. Plants documented as probable diet components include Populus, Platanus, Taxodium, and cycads. Neck posture and skull morphology indicate the animal browsed primarily between 0.5 and 1.5 meters height, and could knock down shrubs or higher branches using its horns and robust skull.
Behavior and senses
Triceratops behavior is reconstructed from bone pathology analyses, horn and frill morphology, and comparison with related ceratopsids. The horns and frill were likely used primarily for intraspecific recognition, sexual display, and dominance fights between males, analogous to modern cervid behavior (Farke et al., 2009). Triceratops skulls frequently show injury marks consistent with impacts from conspecific horns. There is no conclusive evidence of obligate gregarious behavior, although several nearby fóssils suggest social tolerance. The hypothesis that horns were used in defense against T. rex is supported by bite marks on Triceratops fóssils and biomechanical analyses of horn resistance. The ontogenetic pattern of horns and frill changes dramatically from juvenile to adult, which initially led to multiple species being named for the same animal at different growth stages.
Physiology and growth
Triceratops physiology reflects adaptations of a high-energy-demand megaherbivore. Bone histology indicates rapid growth rates in the juvenile phase, consistent with endothermic or near-endothermic metabolism, similar to other studied ceratopsids (Farke, 2011). The bony frill was structurally robust but featured fenestrae (epoccipital openings in some related species), reducing weight without compromising structural integrity for display. Frill vascularization is documented in well-preserved specimens, suggesting a thermoregulatory and in-life color-change signaling role (Hieronymus et al., 2009). The masticatory system was one of the most complex biomechanical structures among dinosaurs: the dental battery combined with robust jaws and powerful temporal and pterygoid musculature to process tonnes of plant matter per year.
Paleogeography
Continental configuration

Scotese & Vérard · CC BY 4.0 · Cretaceous, ~105 Ma
Fossil sites

DBCLS / Togopic, CC BY 4.0
During the Maastrichtiano (~68–66 Ma), Triceratops horridus inhabited Laramidia, the western half of present-day North America, separated from the east by the Western Interior Seaway, a shallow sea dividing the continent. The continents were in very different positions: India was drifting toward Asia, Antarctica was still connected to Australia, and South America was an isolated island.
Bone Inventory
Triceratops is known from dozens of specimens, primarily skulls. Between 1889 and 1891, John Bell Hatcher alone collected over 31 skulls for Marsh. The most complete specimen with well-preserved postcranial skeleton includes sufficient material for substantial reconstruction. Many specimens come from the Hell Creek Formation of Montana and North Dakota.

Found elements
Inferred elements
Scientific Literature
15 papers in chronological order — from the original description to recent research.
Notice of gigantic horned Dinosauria from the Cretaceous
Marsh, O.C. · American Journal of Science
Founding paper in which Marsh describes Triceratops horridus based on horn fragments collected by John Bell Hatcher from the Lance Formation of Wyoming. Marsh initially identified the horns as belonging to a giant Pleistocene bison; after receiving more material, he described the new genus and species of ceratopsian dinosaur, establishing the diagnostic characters of the three-horned skull and bony frill that define the genus.


The Ceratopsia
Hatcher, J.B., Marsh, O.C. & Lull, R.S. · United States Geological Survey Monographs
Monumental 300-page monograph synthesizing all available knowledge on ceratopsians through 1907. Hatcher, Marsh, and Lull describe in detail the anatomy of Triceratops and all ceratopsians then known, establishing the taxonomic and morphological framework that dominated ceratopsian paleontology for decades. The work includes detailed plates of all major specimens and remains an essential reference.


Species resolution in Triceratops: cladistic and morphometric approaches
Forster, C.A. · Journal of Vertebrate Paleontology
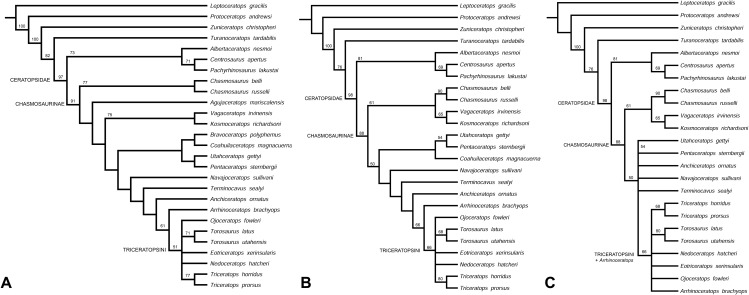
Forster applies cladistic and morphometric analyses to the extensive Triceratops material to resolve the number of valid species. The study concludes that two distinct morphotypes are recognizable: T. horridus, with a longer nasal horn and wider frill, and T. prorsus, with larger supraorbital horns. The work provides the taxonomic foundation on which subsequent ontogenetic studies were built.
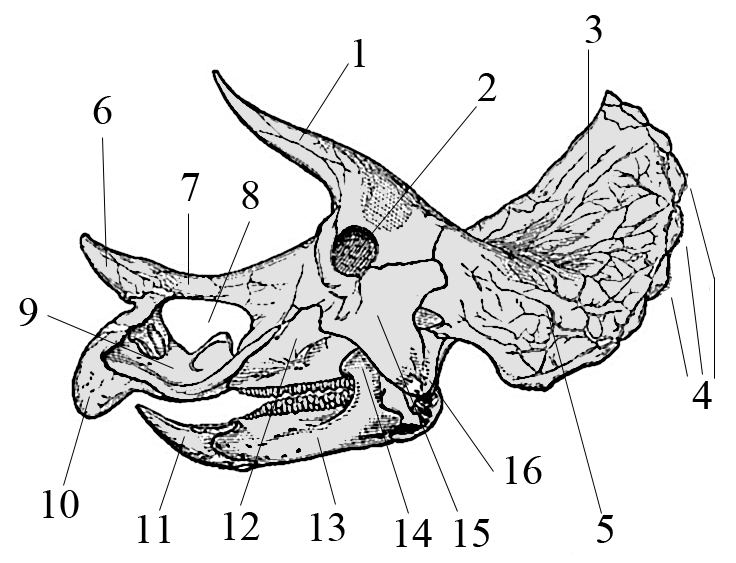

Major cranial changes during Triceratops ontogeny
Horner, J.R. & Goodwin, M.B. · Proceedings of the Royal Society B
Horner and Goodwin demonstrate that the Triceratops skull undergoes radical transformations during ontogenetic development: juveniles have backward-pointing horns that reverse forward with growth. The work documented how ontogenetic changes led earlier paleontologists to create several different species for the same animal at different ages, rationalizing the synonymy of dozens of nominal ceratopsian species.




The facial integument of centrosaurine ceratopsids: morphological and histological correlates of novel skin structures
Hieronymus, T.L., Witmer, L.M., Tanke, D.H. & Currie, P.J. · The Anatômical Record
Hieronymus and colleagues analyze morphological and histological correlates of facial integument in centrosaurine ceratopsids, with relevant comparisons for Triceratops. The study documents evidence that the frill was covered by vascularized skin capable of color change, similar to display structures in modern lizards. The implications for Triceratops display behavior and visual commúnication are significant for modern behavioral reconstructions.


Evidence of combat in Triceratops
Farke, A.A., Wolff, E.D.S. & Tanke, D.H. · PLOS ONE
Farke and colleagues analyze a series of Triceratops skulls for healed bone injuries and identify a pattern consistent with intraspecific combat. Injury frequency in supraorbital horn impact zones is significantly higher than elsewhere on the skull, supporting the hypothesis of male combat for dominance or access to females, analogous to the behavior of modern cervids.


Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny
Scannella, J.B. & Horner, J.R. · Journal of Vertebrate Paleontology
Scannella and Horner propose that Torosaurus, a ceratopsian with a fenestrated frill, was actually the adult form of Triceratops, with the frill becoming fenestrated in very old individuals. The hypothesis was highly controversial and prompted numerous published responses. The debate generated significant methodological advances in determining skeletal maturity in dinosaurs and understanding Late Cretaceous ceratopsian ontogeny.


Anatomy and taxonomic status of the chasmosaurine ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A.
Farke, A.A. · PLOS ONE
Farke redescribes the specimen of Nedoceratops hatcheri, formerly called Diceratops, and evaluates its taxonomic status relative to Triceratops and Torosaurus. The study is relevant to the debate on ontogeny and diversity of chasmosaurine ceratopsids at the end of the Cretaceous. Cladistic analysis positions Nedoceratops as a distinct taxon, not an ontogenetic form of Triceratops, helping resolve part of the controversy.


Torosaurus is not Triceratops: ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy
Longrich, N.R. & Field, D.J. · PLOS ONE
Longrich and Field directly rebut Scannella and Horner's synonymy hypothesis, demonstrating that Torosaurus and Triceratops are distinct genera. The authors identify mature individuals of both genera, show that ventral frill depressions in Triceratops differ in shape and position from Torosaurus parietal fenestrae, and conclude the structures are not intermediates. The debate drove development of more rigorous skeletal maturity criteria for dinosaurs.


Is Torosaurus Triceratops? Geometric morphometric evidence of Late Maastrichtian ceratopsid dinosaurs
Maiorino, L., Farke, A.A., Kotsakis, T. & Piras, P. · PLOS ONE
Maiorino and colleagues apply geometric morphometrics to 28 skulls and 36 squamosals to test whether Torosaurus represents the adult form of Triceratops. The morphospaces of both genera are well separated with distinct ontogenetic trajectories, and the study concludes that Torosaurus is a valid independent taxon. The work provides robust quantitative evidence to rebut the synonymy proposed by Scannella and Horner (2010).


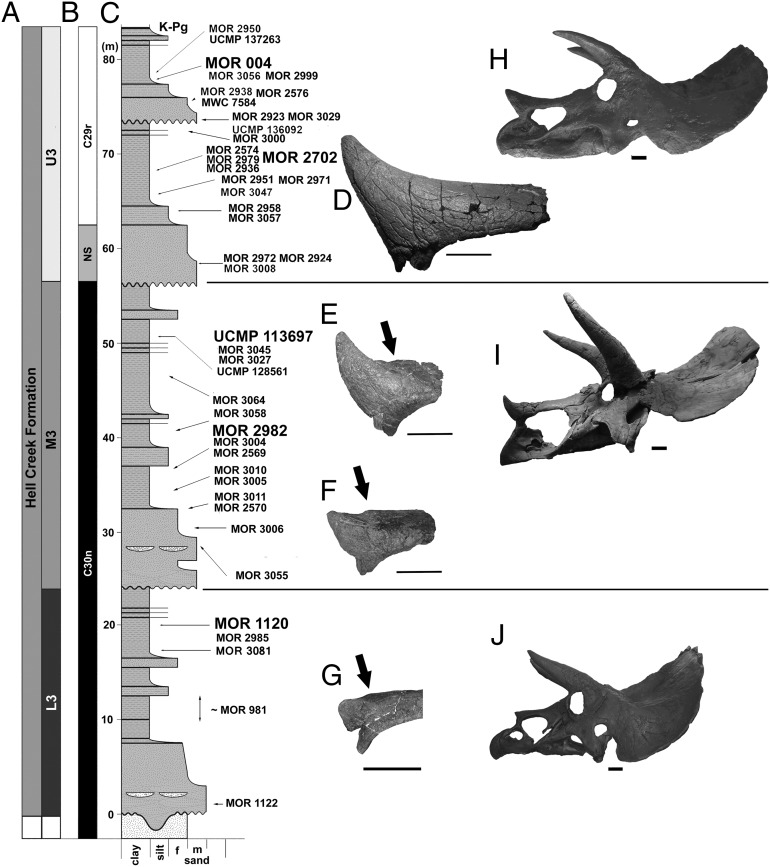
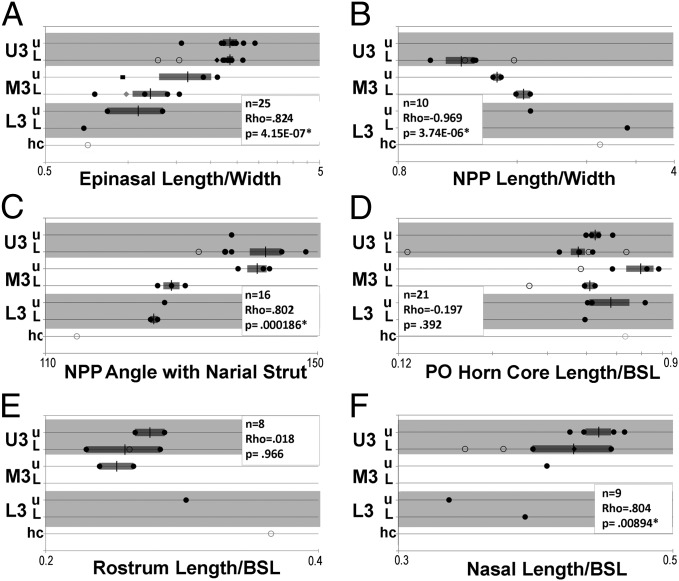
Evolutionary trends in Triceratops from the Hell Creek Formation, Montana
Scannella, J.B., Fowler, D.W., Goodwin, M.B. & Horner, J.R. · Proceedings of the National Academy of Sciences
Scannella and colleagues examine more than 50 Triceratops skulls positioned within the stratigraphic framework of the Hell Creek Formation in Montana. Results show that T. horridus and T. prorsus occupy distinct stratigraphic levels and are connected by transitional morphologies in the middle unit, supporting anagenetic evolution within the lineage. The study rules out sexual or ontogenetic variation as explanations, reinforcing the validity of only two morphologically coherent species.


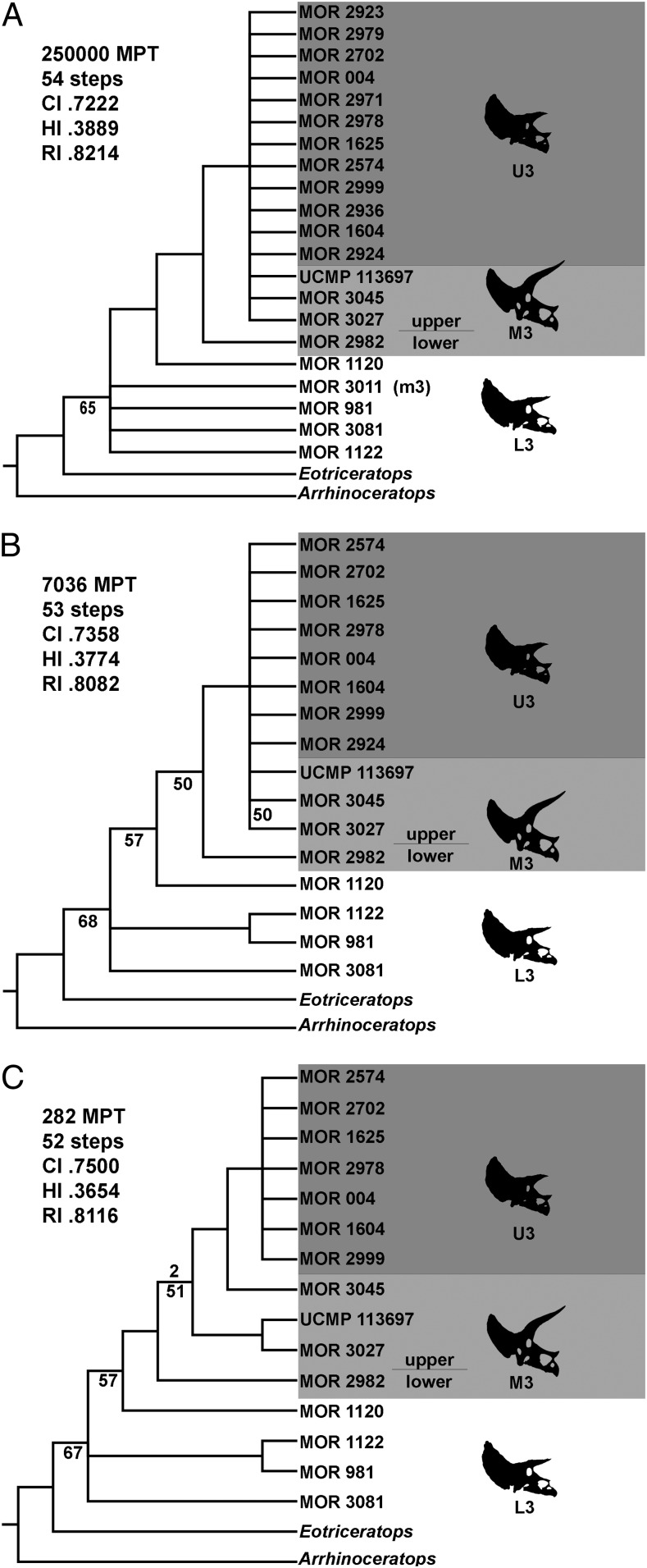
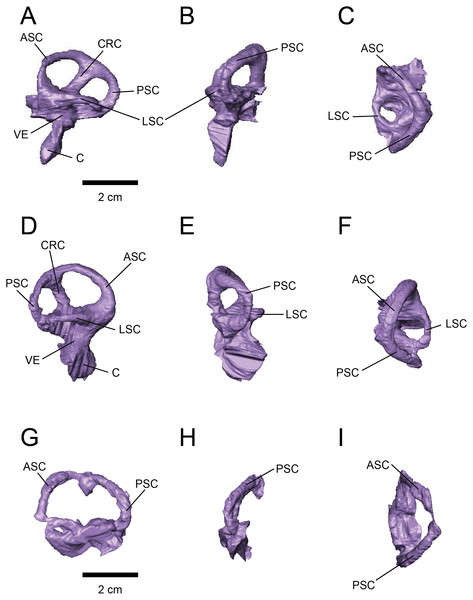
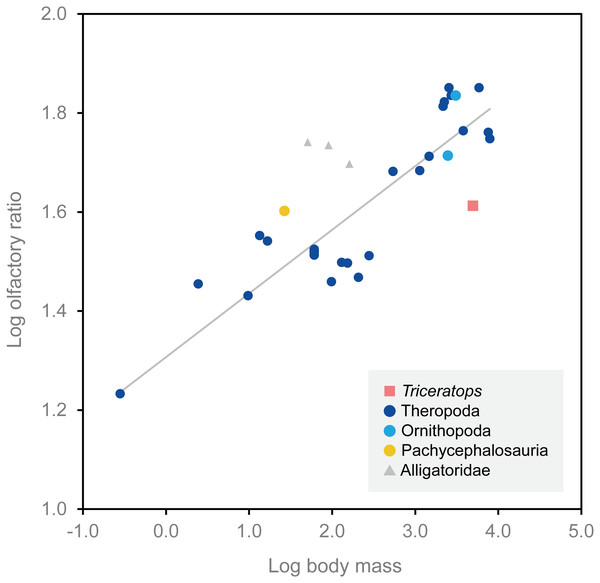
Endocranial anatomy of the ceratopsid dinosaur Triceratops and interpretations of sensory and motor function
Sakagami, R. & Kawabe, S. · PeerJ
Sakagami and Kawabe analyze two Triceratops braincases by computed tomography to produce virtual endocasts of the brain and inner ear. Results indicate that the olfactory bulb was relatively small, suggesting reduced olfaction; the lateral semicircular canal indicates a head posture of approximately 45 degrees, advantageous for displaying horns and frill; and cochlear length suggests adaptation for low-frequency hearing.


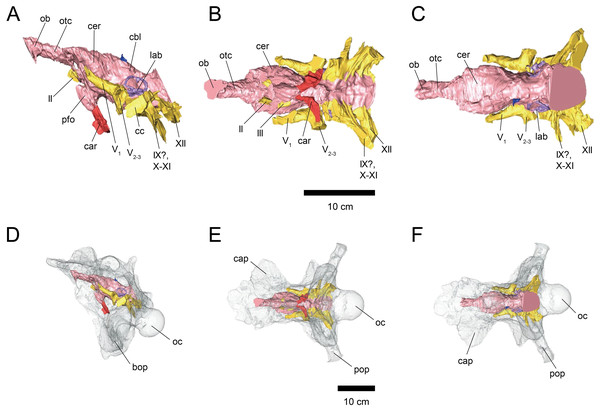
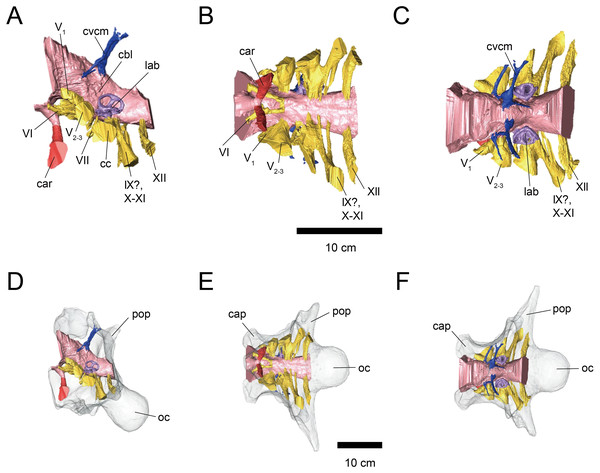
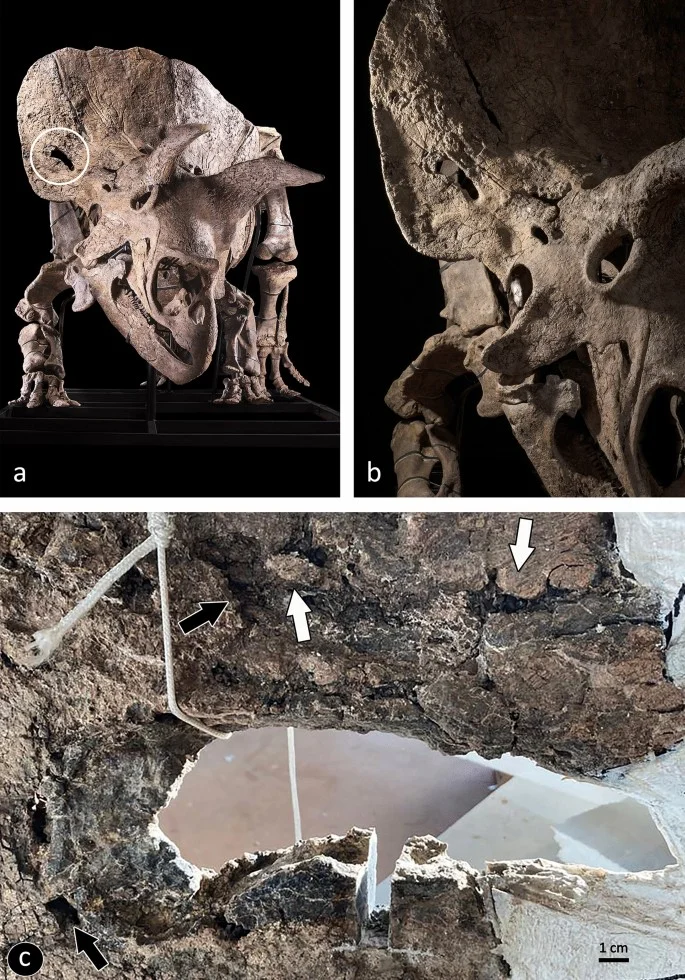
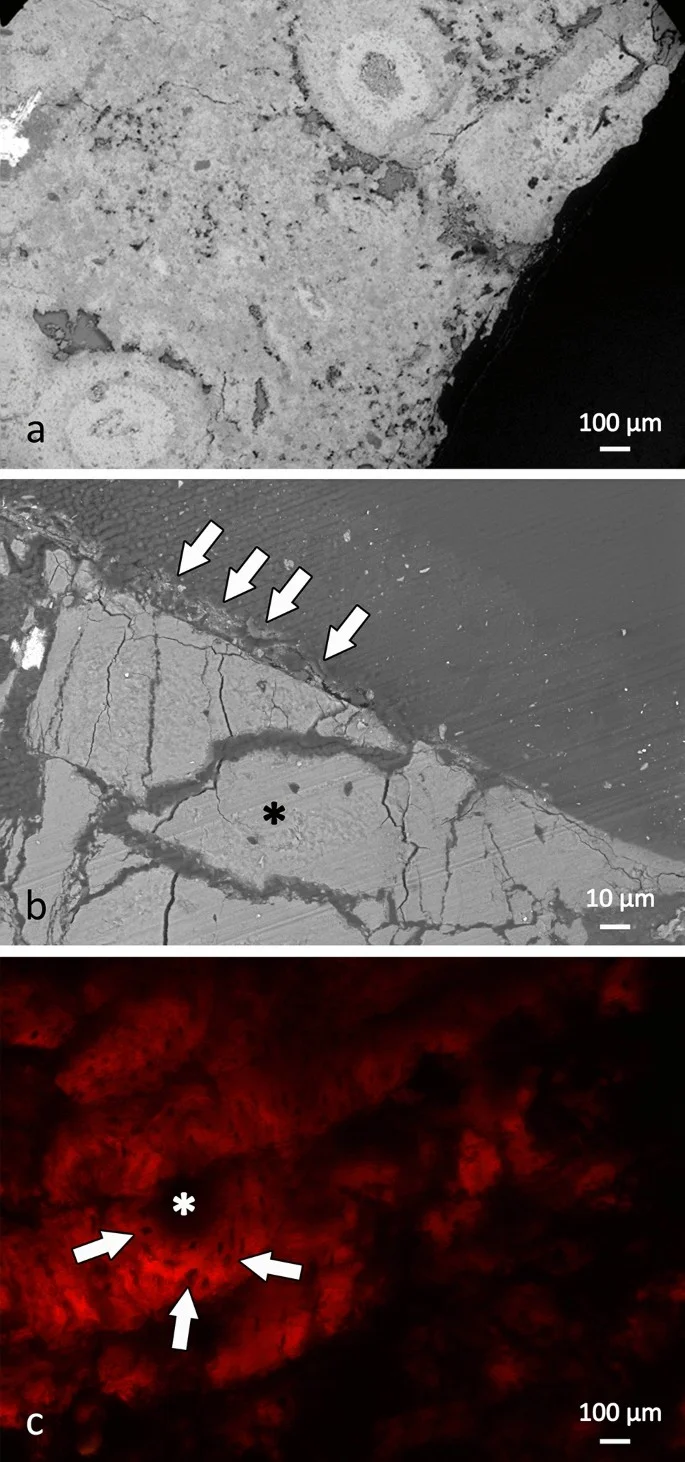
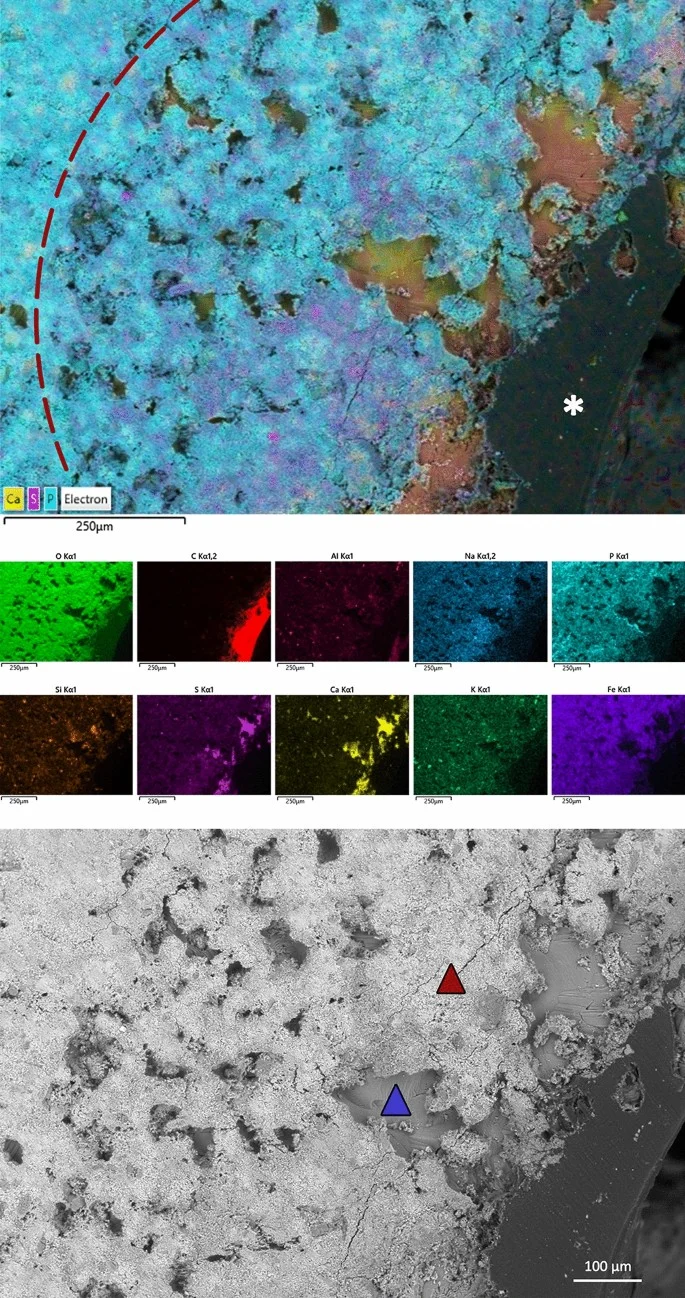
Histological and chemical diagnosis of a combat lesion in Triceratops
D'Anastasio, R., Cilli, J., Bacchia, F., Fanti, F., Gobbo, G. & Capasso, L. · Scientific Reports
D'Anastasio and colleagues microscopically and chemically analyze a large fenestra in the squamosal bone of the Triceratops horridus specimen known as Big John. Histological analysis reveals reactive bone formation and lytic lesions characteristic of the bone remodeling phase. Detected sulfur compounds are consistent with healing bone tissue containing glycosaminoglycans, confirming the lesion as a traumatic wound inflicted during combat with another Triceratops.



How Triceratops got its face: An update on the functional evolution of the ceratopsian head
Nabavizadeh, A. · The Anatômical Record
Nabavizadeh presents a comprehensive review of ceratopsian cranial functional morphology, with special focus on Triceratops. The work integrates research on horn and frill function for combat and display, beak and snout structure, dental wear patterns, cranial musculature, and feeding biomechanics. Results reveal unique masticatory adaptations in Triceratops not seen in other large herbivores, including a powerful and specialized chewing system.


An osteohistological analysis of Triceratops (Ornithischia: Ceratopsidae) cranial ornamentation
Obuszewski, K.D., Smith, N.A. & Brown, G.R. · The Anatômical Record
Obuszewski and colleagues histologically analyze cranial ornamentation of a subadult Triceratops specimen, documenting previously undescribed vascularization patterns in the postorbital horns. The squamosal exhibits a novel ontogenetic shift from rapid azonal to slow zonal growth late in development. The study proposes that occipital condyle dimensions can distinguish older from younger individuals, with implications for ceratopsid taxonomy.


Classification
Discovery
Fun fact
Triceratops has the largest skull relative to body size of any known terrestrial animal: the skull reached 2.5 meters, equivalent to one third of the total body length. This makes it a record holder among all known terrestrial vertebrates.
Last reviewed: April 24, 2026