Paleobiology
Dinosaur Biology
How did the largest land animals in history work? From metabolism to social behavior, from feathers to intelligence: what fossil evidence reveals about dinosaur biology.
Why did they get so big?
Brachiosaurus weighed 50 tons. Patagotitan, discovered in 2017, may have reached 70. To understand how land animals grew to such sizes, you need to look at anatomy.
🫁
Respiratory system with air sacs
Birds breathe unidirectionally: air passes through the lungs in one direction only, never "going back." Dinosaurs did the same, using air sacs that worked like bellows. This extracts far more oxygen per breath than the bidirectional system of mammals, making large bodies energetically viable. A 50-ton body with mammalian lungs would be impossible: it could not oxygenate tissues efficiently enough.
🦴
Hollow bones
Brachiosaurus vertebrae were up to 60% air. Sauropod bones had internal chambers connected to the air sacs, maintaining a rigid structure without the corresponding weight. A 6-ton African elephant has proportionally denser bones than a 50-ton sauropod. This skeletal pneumatization is one of the reasons dinosaurs and birds share the ability to grow beyond mammalian limits.
🌿
Swallowing without chewing
Sauropods did not chew. They tore off plants and swallowed them whole, letting gut fermentation do the work. A small, lightweight head atop an enormous neck consumed vegetation in large volumes without wasting energy on chewing. The smaller the head, the easier it was to support the long neck. It is the opposite of an elephant, which has a massive head full of chewing muscles and high-crowned molars.
📈
Rapid growth like birds
Reptiles grow slowly throughout their lives. Birds grow fast and stop. Dinosaurs grew like birds. A young sauropod gained over 2 tons per year at peak growth rate and reached adult size in 20 to 30 years. This reduced the time the animal remained vulnerable to predators. An elephant takes 20 years to reach 5 tons. Dinosaurs reached 10 times that in a similar timeframe.
🌳
Abundant plant life in a warm world
The elevated CO₂ of the Mesozoic was fuel for plants. Dense forests covered latitudes that today are deserts or tundra. More plant biomass meant giant herbivores could feed without depleting their environment. A sauropod needed to eat hundreds of kilograms of vegetation per day. This was only possible because food was everywhere.
💨
The role of oxygen
Atmospheric O₂ rose to about 30% in the Late Cretaceous, compared to 21% today. This helped, but there is important context: the giant insects of the Carboniferous, 300 million years before the dinosaurs, grew with O₂ at 35%. They were large for completely different reasons. For dinosaurs, the air sac system is the central respiratory factor. Oxygen levels were a facilitator, not the cause.
Did cold climate make dinosaurs grow bigger?
It is common to see the idea that climatic cooling at the end of the Cretaceous pushed dinosaurs toward giant sizes, applying Bergmann's rule (warm-blooded animals get larger in cold climates to conserve heat). A 2025 study tested this hypothesis for tyrannosaurs with rigorous statistics and arrived at a more nuanced answer.
The study mapped three distinct variables onto the same phylogenetic tree of tyrannosauroids: body mass, body length, and mean annual temperature of the location where each species lived. Geological time runs from left (Jurassic, 200 million years ago) to right (end of the Cretaceous, 66 million years ago). Branch colors indicate the estimated value of the variable at that evolutionary moment: cool tones (purple, blue) are low values, warm tones (yellow, orange, red) are high values. Compare the three panels below.
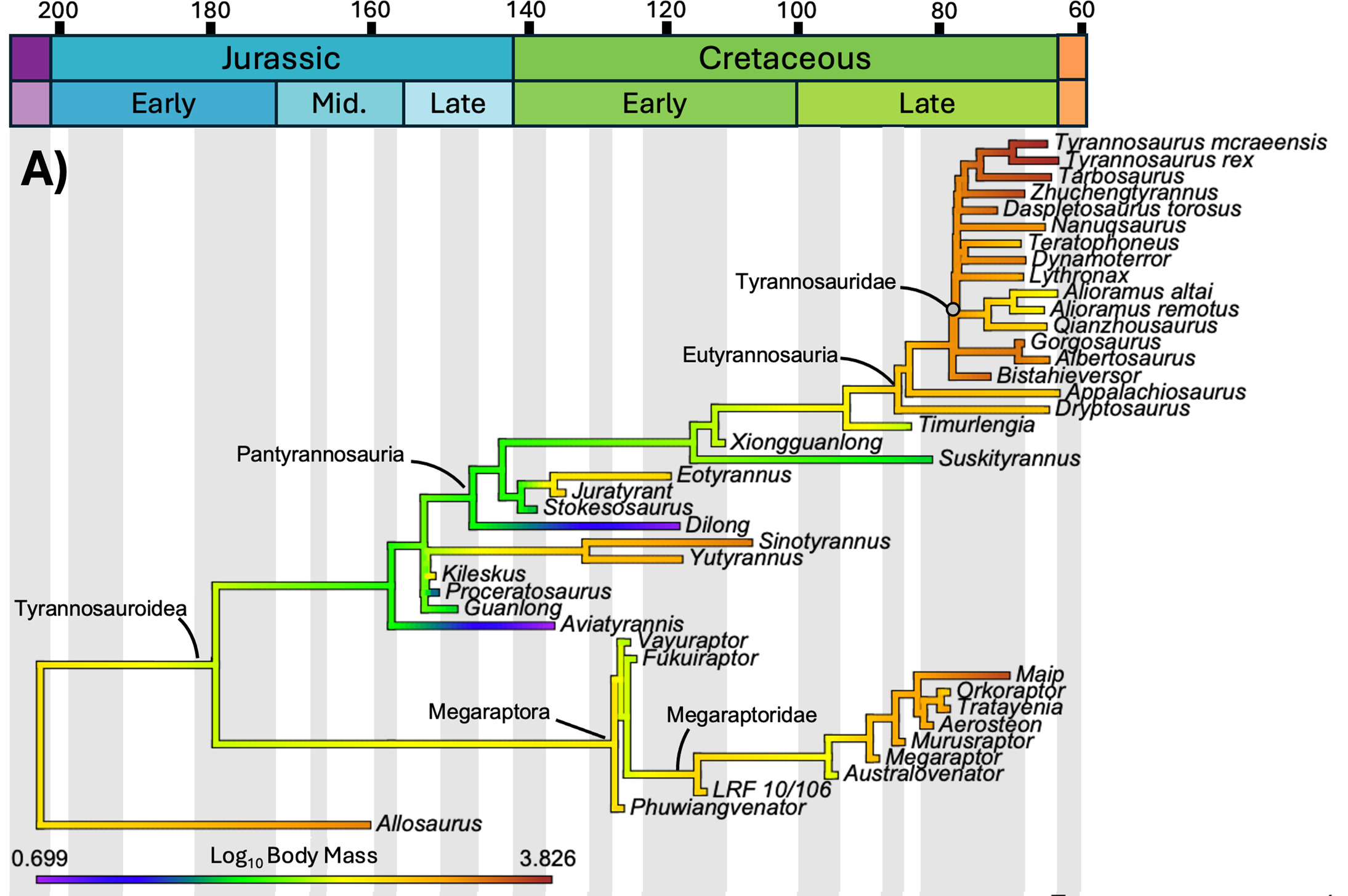
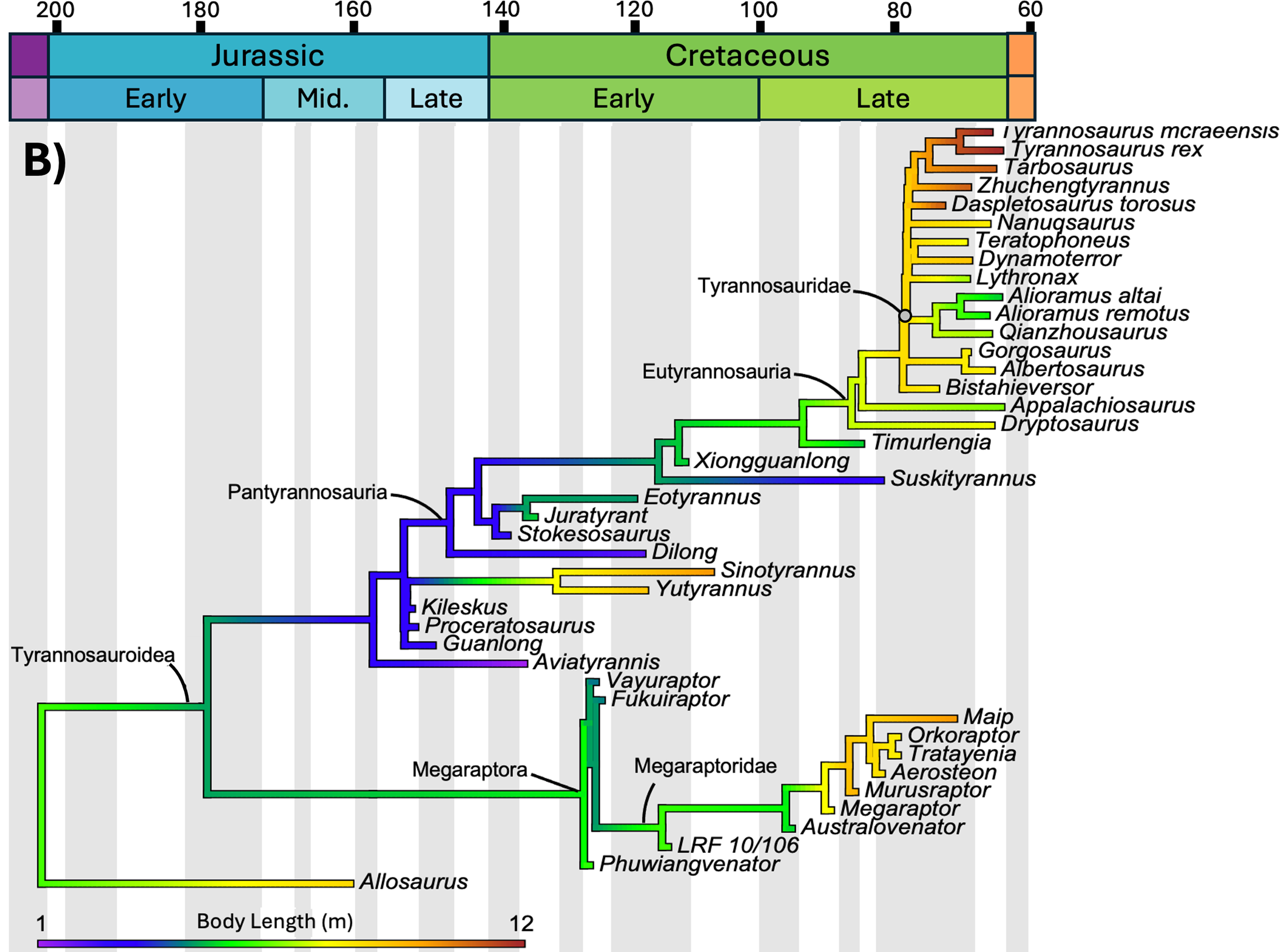
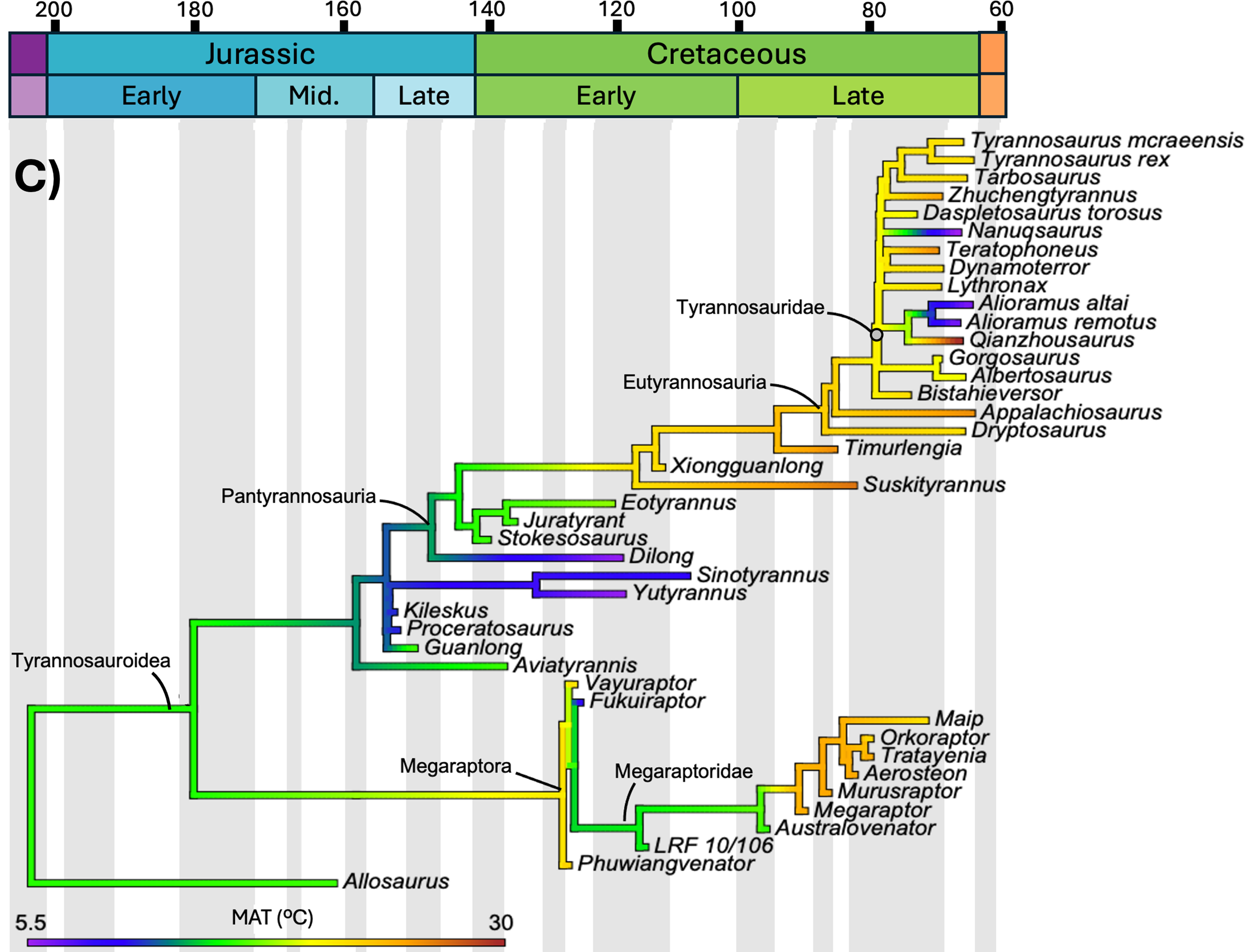
The statistical test
Morrison and colleagues mapped the evolution of body mass and length of every known tyrannosaur against the mean annual temperature (MAT) of its time and region. The correlation between temperature and body mass was statistically non-significant: R² of 0.303 and p of 0.051, right at the threshold of significance but not crossing it. Even after log-transforming the data, the result did not improve (R² of 0.293, p of 0.060).
In plain language: temperature does not predict the body size of a tyrannosaur. There is no direct relationship of the "it got colder, they got bigger" type.
But the study also shows that tyrannosaurs really did become gigantic at a specific moment: right after the so-called Cretaceous Thermal Maximum, between 90 and 95 million years ago, when global temperatures began to fall toward the Campanian and Maastrichtian. The authors' answer to this apparent contradiction is that climate acted indirectly.
Post-Thermal Maximum cooling contributed to the extinction of the carcharodontosaurids, the apex predator group that included Giganotosaurus, Carcharodontosaurus, and Mapusaurus. These animals were 12 to 13 meters long and dominated the top of the food chain across most of the world until the end of the Cenomanian. When they disappeared, the ecological slot of "colossal predator of very large prey" was left open. That is when the tyrannosaurines, until then mid-sized carnivores of 5 to 7 meters, grew in just a few million years to the 12 meters of T. rex. In other words, climate did not make tyrannosaurs bigger directly; it killed the competition and opened up the niche.
A curious detail reinforces the reasoning: Yutyrannus huali, a basal tyrannosaur of 9 meters that lived in China 125 million years ago, in a cold, high-altitude environment, was already gigantic for its time. The authors suggest that the capacity to reach large size in colder climates may be an ancestral feature of the lineage, only realized at scale when the niche allowed. But they caution that the pattern is not uniform across the tyrannosaur fossil record and needs more data to be confirmed.
Source: Morrison, C., Pittman, M., Nurgaliyev, B., & Salguero-Gómez, R. (2025). Rise of the king: Gondwanan origins and evolution of megaraptoran dinosaurs. Royal Society Open Science, 12(5), 242238. doi.org/10.1098/rsos.242238
Did warm climate make dinosaurs grow bigger?
The symmetric question also needs to be asked. If cold does not make dinosaurs grow, does heat? The mid-Cretaceous was the warmest period of the entire Mesozoic Era and also housed the largest land animals that ever walked the planet. Cause and effect, or coincidence?
A world on fire (literally)
The Cretaceous Thermal Maximum occurred during the Cenomanian-Turonian interval, peaking around 94 million years ago. It was the warmest period of the last 145 million years: there was no ice at the poles, temperate forests reached the north of Alaska and Antarctica, and sea surface temperatures in the tropics and mid-latitudes exceeded 35 °C, a value recorded today only in the Persian Gulf at the peak of summer.
The 485-million-year reconstruction by Judd, Tierney, Lunt and colleagues (Science, 2024) confirms the scale: global mean surface temperature (GMST) at the Cretaceous peak rose to about 35 to 36 °C, against the current average of 14 to 15 °C. That is, the entire planet was, on average, 20 °C warmer than it is today. The long Phanerozoic marine δ¹⁸O curve shows the same signal: the Cretaceous is one of the three great heat peaks of Earth history, alongside the Devonian and the Early Triassic.
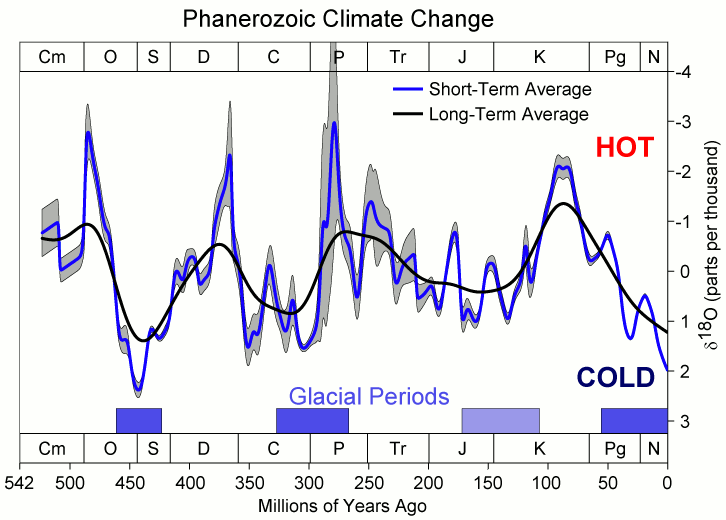
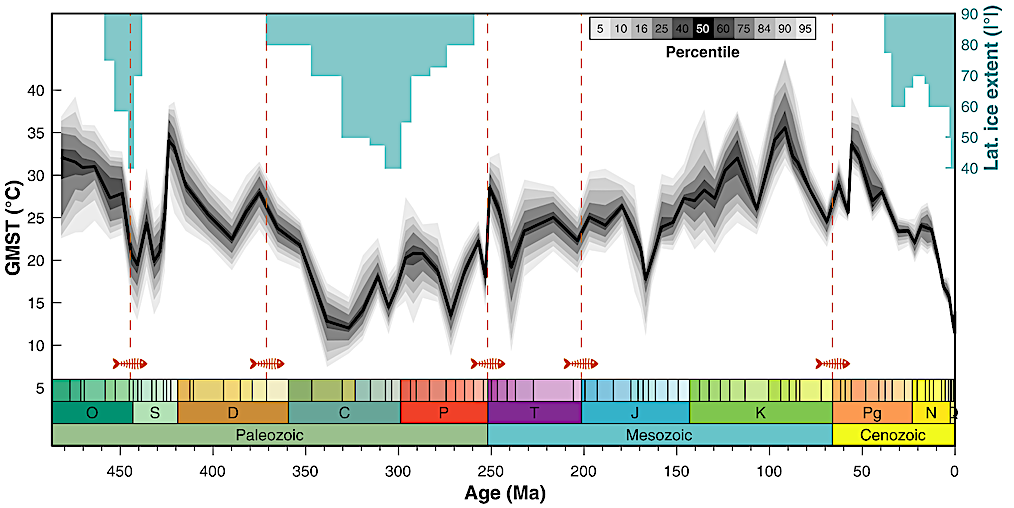
After this peak, around 90 to 85 million years ago, global temperatures began to fall slowly. The drop continued through the Campanian (83 to 72 Ma) and the Maastrichtian (72 to 66 Ma), the last two stages of the Cretaceous, the world in which T. rex, Tarbosaurus, and Triceratops lived. These terminal dinosaurs therefore inhabited a Cretaceous "already cooling" relative to the thermal peak, but still with GMST about 6 to 8 °C above the current average, according to the same Judd et al. reconstruction.
The question then becomes: where were the largest dinosaurs, at the peak of the heat or later, when it cooled?
The answer, taking the fossil record seriously, is direct: the largest known dinosaurs lived precisely at the peak of the heat. Argentinosaurus huinculensis, a titanosaur from Patagonia, has been dated to 97 to 93 million years ago, squarely in the Thermal Maximum zone, with mass estimates ranging from 65 to 90 tonnes, probably the largest land animal ever catalogued. Patagotitan mayorum, another Argentine titanosaur, comes from layers around 101 million years old, just before the peak, at 55 to 77 tonnes. Among the carnivores of the same interval are Giganotosaurus, Mapusaurus, the North African Carcharodontosaurus, and Spinosaurus aegyptiacus, all 12 to 15 meters long.
So does the warm mid-Cretaceous beat the cool terminal Cretaceous in gigantism? In the sauropod lineage, yes. The "largest land animal" crown belongs to the Cenomanian. A 90-tonne titanosaur is almost ten times heavier than a T. rex. In pure mass terms, the peak of gigantism in the Kingdom of the Dinosaurs was in the warmest period, not the coldest.
But that does not mean heat caused gigantism directly. The consensus interpretation is that the Thermal Maximum created ecosystems of extreme ecological productivity: tropical forests extending across almost the whole planet, shallow, warm epicontinental seas irrigating the continents, plants growing year-round, strong insolation all the time. This gigantic plant biomass sustained herds of 60- to 90-tonne titanosaurs, which in turn sustained 12- to 15-meter predators. It was the climate that made such an ecosystem possible, not heat acting on the animals' bodies.
The proof lies in what came next. When the planet began to cool at the end of the Cenomanian-Turonian, the largest Gondwanan titanosaurs and the carcharodontosaurids went extinct. Some groups shrank, others disappeared from the record. But gigantism did not vanish: the tyrannosaurines (T. rex, Tarbosaurus) and some Laurasian titanosaurs such as Alamosaurus filled the open niches and reached remarkable sizes in the Campanian-Maastrichtian, in a cooler climate. Just on a smaller scale than the giants of the thermal peak.
Direct answer
The largest dinosaurs of all time lived at the peak of the heat (Cenomanian-Turonian, 100 to 90 million years ago). But it was not the heat that made them grow; it was the ecological productivity that the warm, humid climate made possible. When it cooled, some giants went extinct and others appeared (T. rex and company), but none came close to the 70 to 90 tonnes of the Cenomanian titanosaurs. Neither cold nor heat "makes" dinosaurs grow directly. Climate always acts through the shortcuts of plant productivity, the size of prey herds, and the ecological slot left by those that go extinct.
Sources: chronological and mass data for Argentinosaurus and Patagotitan compiled from Wikipedia/Argentinosaurus and Wikipedia/Patagotitan, based on Benson et al. 2014, Carballido et al. 2017, and Paul 2019. Cretaceous Thermal Maximum temperatures: Judd, Tierney, Lunt et al. 2024, Science 385, eadk3705, with complementary data from Huber et al. 2018. Phanerozoic δ¹⁸O curve adapted from Veizer et al. 1999 and Prokoph et al. 2008. Synchronized extinction of carcharodontosaurids and diplodocoids at the Cenomanian-Turonian discussed in Novas et al. 2005 and Sales et al. 2016.
How big were they, exactly?
Numbers in meters and tonnes are abstract. A human silhouette next to the animal turns those values into immediate scale. The carousel below brings together 25 species from the Jurassic and the Cretaceous, with length and weight estimated from the literature and the visual comparisons most commonly used in scientific papers.
Warm or cold blooded?
For decades, paleontologists debated whether dinosaurs were ectothermic (cold-blooded), like reptiles, or endothermic (warm-blooded), like birds and mammals. The answer lies in between, and the path to that answer is one of the most fascinating stories in modern paleontology.
The historical debate
Until the 1960s, the consensus was that dinosaurs were giant cold-blooded lizards: slow, sun-dependent, limited by ambient temperatures. In 1968, Robert Bakker published a paper that inaugurated the so-called "Dinosaur Renaissance," arguing that upright posture, rapid growth rates, and predator-prey ratios in Mesozoic ecosystems were incompatible with ectothermy. Bakker argued that dinosaurs were as endothermic as mammals. The traditional view, championed by researchers like R. Tracy and James Farlow, argued that ectothermy would explain the energy efficiency needed to sustain such large bodies. The reality, as almost always in biology, turned out to be more complex than either extreme.
What the bones reveal
The widespread presence of fibrolamellar bone in dinosaurs indicates rapid, continuous growth. This type of bone tissue forms when an animal grows so fast that the osteocytes (bone cells) get "trapped" in a disorganized matrix, without time to arrange themselves in concentric layers as in the lamellar bone of reptiles. It is the same type of bone found in birds and mammals. At the same time, many species show Lines of Arrested Growth (LAGs), seasonal pauses in growth that form visible rings in thin bone sections, comparable to tree rings. These pauses are common in ectotherms, which stop growing during cold or dry seasons. This paradoxical combination, rapid growth with seasonal pauses, led Grady et al. (2014) to propose the concept of mesothermy: an intermediate metabolism between reptiles and mammals, after analyzing growth rates of 21 dinosaur species compared to over 360 living species. According to the study, dinosaurs had metabolic rates higher than any living reptile, but lower than most mammals.
Isotope thermometry
Geochemical studies brought independent evidence. Eagle et al. (2011) used clumped isotope thermometry on fossilized eggshell carbonates. The principle is elegant: the frequency with which heavy isotopes of carbon-13 and oxygen-18 bond within the same carbonate ion depends on the temperature at which the mineral formed. The colder the temperature, the greater the "preference" for these heavy isotopes to cluster together. Analyzing sauropod eggshells from the Allen Formation (Patagonia, Argentina), Eagle estimated body temperatures of 36 to 38°C, values close to modern mammals (37°C) and well above typical ectothermic reptiles (which track ambient temperature). Dawson et al. (2020) expanded this approach, analyzing oxygen isotopes in eggshells from both major dinosaur lineages (Ornithischia and Saurischia), concluding that elevated body temperatures relative to the environment were likely an ancestral trait of the entire group, present before the diversification of dinosaurs.
Variation across groups
Thermal physiology varied enormously within Dinosauria. Small theropods, ancestors of birds, were likely fully endothermic, with thermal insulation provided by feathers and metabolic rates high enough to maintain constant body temperature regardless of environment. Large sauropods may have maintained stable temperatures through a completely different mechanism: gigantothermy (inertial homeothermy). Enormous body mass works as a thermal reservoir: a 30-ton animal has such a low surface-to-volume ratio that it takes days to lose significant heat, even without active metabolic heat generation. This phenomenon is observed today in leatherback sea turtles (Dermochelys coriacea), which maintain core temperature 8-10°C above surrounding water despite being ectothermic. For medium-sized dinosaurs, such as ornithopods of 500 kg to 2 tons, the situation was more ambiguous: they probably generated active metabolic heat, but less efficiently than mammals. The simplified division between "warm-blooded" and "cold-blooded" is inadequate to describe the physiological diversity of these animals.
Nasal turbinates: the clue that didn't preserve
Mammals and birds possess thin bony structures inside the nasal cavity called respiratory turbinates. These structures warm and humidify inspired air and recover water from exhaled air, essential for endothermic animals that lose substantial water through rapid breathing. Ruben et al. (1996) argued that the absence of turbinates in dinosaurs indicated ectothermy. The problem is that these structures are extremely fragile and rarely preserve in fossils, even in mammals. CT scans of dinosaur skulls reveal complex nasal cavities with ridges and grooves that could have supported equivalent soft tissue, but the evidence remains inconclusive. Absence of evidence, in this case, is not evidence of absence.



Feathers or scales?
The discovery of feathered dinosaurs in Liaoning, China, from the 1990s onward, revolutionized paleontology. Feathers were not exclusive to birds, and their original functions had nothing to do with flying.
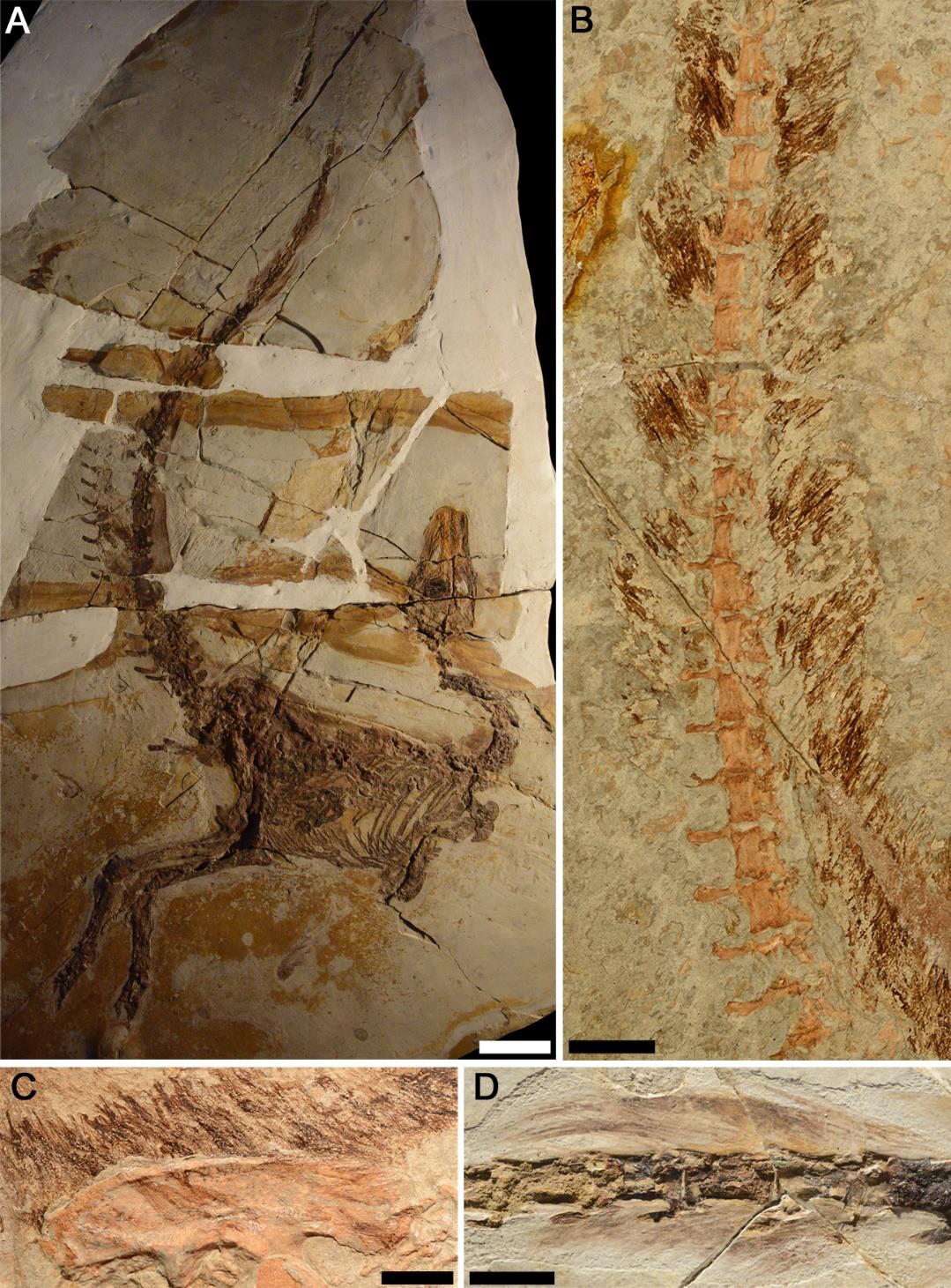
The Liaoning revolution
In 1996, Sinosauropteryx was described as the first non-avian dinosaur with feather-like integumentary structures (proto-feathers). The fossil, preserved in ultra-fine-grained limestone of the Yixian Formation, clearly showed a halo of filaments around the body, including countershading patterns (dark dorsum, light belly) indicating functional coloration. Since then, dozens of feathered species have been found in the volcanic deposits of Liaoning, which preserve soft tissues with extraordinary detail thanks to rapid burial by volcanic ash in shallow lakes. This type of geological deposit is called a Lagerstatte (German for "storage place"), and Liaoning is considered the most important in the world for feathered dinosaurs.
Key species
Microraptor gui (2003) is a dromaeosaurid with four "wings," asymmetrical flight feathers on both fore and hind limbs, evidencing gliding or even rudimentary flight capability. Aerodynamic analyses suggest it glided between trees, using the rear feathers as stabilizers. Anchiornis huxleyi (2009) preserved melanosomes (pigment-containing organelles) so well that researchers reconstructed its complete coloration: dark gray body, red crest, white flight feathers with black tips. It was the first dinosaur with full-body color mapping. Yutyrannus huali (2012) is the largest known feathered dinosaur: a tyrannosauroid of ~9 meters and ~1.4 tons with filaments covering much of its body. Its existence demonstrated that feathers were not exclusive to small dinosaurs.
Beyond theropods
In 2014, Kulindadromeus zabaikalicus, found in Russia, expanded the picture: it is an ornithischian with feather-like structures, including three distinct types of filamentous coverings. Ornithischians are the opposite lineage to theropods in the dinosaur family tree. The presence of plumose structures in both lineages suggests that some form of filamentous covering may be ancestral to all dinosaurs, arising a single time in the common ancestor of the entire group. If correct, scaly dinosaurs like sauropods and ceratopsians would have lost filamentous structures secondarily, not the other way around. The idea is still debated, but gained strength after the description of Kulindadromeus.
Evolutionary stages of feathers (Prum model)
Richard Prum (1999) proposed a five-stage model for feather evolution, from simplest to most complex: I. Single hollow filament (as in Sinosauropteryx). II. Filaments branching from a base (plumulaceous). III. Barbules with hooks that lock barbs together (planar feathers). IV. Asymmetry between barbs (functional for aerodynamics). V. Further diversification (specialized flight, display feathers, etc.). This model predicted the existence of intermediate-stage feathers years before fossils like Anchiornis and Kulindadromeus confirmed stage I and II structures in non-avian dinosaurs.
The T. rex case: feathers or scales?
T. rex itself is the subject of intense debate. Skin impressions published by Bell et al. (2017) revealed scales on areas like the abdomen, tail, and neck, with textures similar to crocodile skin. This would indicate that large tyrannosaurs may have lost feathers secondarily, possibly due to thermoregulatory pressure: large animals in warm climates need to dissipate heat, and feathers interfere. It is the same principle that explains why elephants and rhinos lost fur. However, Bell's skin impressions cover only ~1% of the body surface, leaving open the possibility that T. rex had feathers in unpreserved regions, such as the back or crest.
Original functions of feathers
Feathers did not evolve for flight. Evidence points to multiple prior functions: sexual display and visual communication (color patterns preserved in fossils, including iridescence in Microraptor, detected by Li et al. in 2012); thermal insulation (particularly important for small endothermic dinosaurs in cold climates); egg protection during brooding (oviraptorids fossilized over nests with arms spread, in a position identical to modern birds covering eggs with feathers); and possibly camouflage (countershading in Sinosauropteryx). Flight came later, as a functional co-option of already existing structures, likely passing through intermediate gliding stages, as seen in Microraptor.
Distribution by group
Feathered
Coelurosaurs (tyrannosauroids, dromaeosaurids, birds)
Scaled
Sauropods, ankylosaurs, ceratopsians (mostly)


Solitary or in herds?
Dinosaurs displayed a wide range of social behaviors: from solitary habits to herds with thousands of individuals. Deciphering the behavior of extinct animals is one of paleontology's greatest challenges, but the fossil record provides surprisingly detailed clues.
Bonebeds: collective graveyards
The strongest evidence for gregarious behavior comes from bonebeds: fossil deposits where hundreds or thousands of individuals of the same species were preserved together. The Hilda bonebed in Alberta, Canada, contains remains of over 1,000 individuals of Centrosaurus apertus spread across 2.3 km², interpreted as an entire herd killed during a flood while trying to cross a river (Eberth & Getty, 2005). The Maiasaura bonebeds in the Two Medicine Formation, Montana, USA, with up to 10,000 estimated individuals, indicate enormous herds that likely migrated seasonally (Varricchio & Horner, 1993). The critical question in bonebed interpretation is taphonomy: how to distinguish between animals that lived together and died simultaneously versus carcasses transported by water and accumulated at the same location over time? The presence of individuals of all age classes (hatchlings to adults) and the absence of size-based sorting in the deposit favor the interpretation of mass death of a cohesive social group.
Trackways and migration
Parallel fossilized trackways of sauropods, such as those at the Purgatoire River in Colorado (over 1,300 footprints across 100 trackways), confirm that groups walked together in the same direction and at the same speed. Analysis of these trackways allows calculation of individual speeds and verification of whether animals moved in a coordinated fashion. At the Cal Orcko site in Bolivia, more than 5,000 footprints from 462 trackways of multiple species cover a limestone wall of 25,000 m², one of the largest records of dinosaur activity in the world. Sauropod trackways at sites like Davenport Ranch, Texas, show footprints of small individuals at the center and large ones at the edges, suggesting protective behavior similar to elephant herds, which position calves in the middle of the group during movement.
Parental care
Parental care is also well documented. Maiasaura peeblesorum, whose name means "good mother lizard," was described by Jack Horner in 1979 from nesting colonies at Egg Mountain, Montana. Nests were spaced about 7 meters apart (approximately the length of the adult animal), with hatchlings that remained in the nest until doubling in size, evidenced by dental wear on the hatchlings (they were already eating, but not yet walking). Oviraptorids like Citipati osmolskae were found fossilized directly on top of their nests in brooding position, arms extended over the eggs like modern birds. In 2021, Pol et al. published the discovery of a nesting colony of Mussaurus patagonicus in Patagonia, Argentina, with 80 eggs and 100 skeletons organized by age class: adults in one sector, juveniles in another, eggs in a third. This suggests age segregation within a social group, unprecedented in the dinosaur fossil record.
Communication
Social life requires communication, and dinosaurs had sophisticated anatomical structures for it. The hollow crest of Parasaurolophus functioned as a resonating chamber, producing low-frequency sounds (~30-700 Hz) for long-distance communication (Weishampel, 1981). Digital reconstructions of the crest's internal tube allowed simulation of the probable sound: a deep bellow, similar to a wind instrument. Shields and horns of ceratopsians like Triceratops likely served for visual species recognition and sexual selection (Padian & Horner, 2011). The enormous variation in shield shape among closely related species (e.g., Triceratops vs. Torosaurus vs. Chasmosaurus) supports this hypothesis. In hadrosaurs, the diversity of crest shapes within the same family suggests each species had its own "sound channel," allowing individuals of different species to coexist without signal confusion.
Theropods: solitary or gregarious?
Most large theropods, like T. rex and Allosaurus, were probably solitary or territorial, similar to modern big cats. However, exceptions exist. Sites with multiple specimens of Mapusaurus in Argentina and Albertosaurus in Alberta suggest some degree of social aggregation in large theropods. Currie (1998) described a bonebed with 12 individuals of Albertosaurus sarcophagus of different ages, which he interpreted as a social group. The interpretation is debated: the animals may have been independently attracted to a carcass or water source, as occurs with modern crocodiles, without real cooperation.


How did predators hunt?
Hunting machines refined by 165 million years of evolution. Each group of predators developed distinct strategies, from the crushing bite of T. rex to the surgical precision of raptors and the aquatic hunting of spinosaurids.
The T. rex bite
T. rex had the most powerful bite ever recorded in a land animal: between 35,000 and 57,000 newtons, the equivalent of nearly 6 tons of force concentrated on a single posterior tooth (Bates & Falkingham, 2012). For comparison, a modern lion generates ~4,000 N and a saltwater crocodile ~16,000 N. This pressure was enough to crush bones and extract marrow, a behavior called osteophagy, confirmed by coprolites (fossilized feces) of T. rex containing crushed bone fragments (Chin et al., 1998). Its serrated teeth (ziphodont dentition) worked like steak knives: the micro-serrations reduced the force needed to cut through flesh, while internal dentine structures reinforced each denticle against fracture. Each tooth was about 15-30 cm long, with deep roots anchored in the jaw by robust periodontal ligaments. Teeth were continuously replaced: when one fell out, another was already growing beneath it, a cycle lasting about 2 years per tooth.
Senses and locomotion
The binocular vision of T. rex covered about 55 degrees (Stevens, 2006), with eyes 13 cm in diameter, larger than those of any living land animal. CT scans of the skull reveal proportionally enormous olfactory bulbs, suggesting a sense of smell comparable to modern vultures, capable of detecting carcasses from kilometers away. The inner ear indicates sensitivity to low-frequency sounds, useful for locating large prey. Definitive proof of active predation, rather than mere scavenging, came in 2013: a T. rex tooth was found lodged between healed vertebrae of a hadrosaur (Edmontosaurus), demonstrating the prey was alive when attacked and survived the assault (DePalma et al., 2013). Despite its size, T. rex was probably not fast: biomechanical estimates indicate 20-29 km/h as maximum sustainable speed (Hutchinson & Garcia, 2002). Above that, the leg bones would suffer stress fractures. Its prey, such as Edmontosaurus, were equally slow, making the relative speed adequate.
Alternative strategies: Allosaurus and Spinosaurus
Not all predators worked like T. rex. Allosaurus, the dominant Jurassic predator, had a weaker jaw but a lightweight skull and powerful neck muscles. Rayfield et al. (2001) proposed the "hatchet bite" hypothesis: Allosaurus opened its mouth to ~80 degrees and drove its skull down onto the prey with force, using the upper jaw as a cutting weapon. The curved, serrated teeth ripped flesh on impact, causing massive hemorrhage. Spinosaurus aegyptiacus, in turn, was a semiaquatic predator. Ibrahim et al. (2014, 2020) demonstrated that its long, conical snout filled with pressure receptors similar to those of crocodiles, and its laterally compressed tail (like a giant salamander's) indicate adaptation for hunting fish and other aquatic animals. The conical, unserrated teeth were ideal for gripping slippery prey, not for cutting flesh.
RPR model: how raptors hunted
Fowler et al. (2011) proposed the Raptor Prey Restraint (RPR) model: dromaeosaurids like Deinonychus leaped onto prey, pinned it with the curved claws of their second toe (the famous "retractable claw"), and consumed it while still alive, flapping their feathered arms for balance as the prey tried to escape. Exactly as eagles and hawks do today when capturing prey larger than themselves. The second-toe claw, popularly believed to be a slashing weapon, actually functioned more as a pinning hook: biomechanical analyses show the tip was optimized for piercing and gripping, not tearing (Manning et al., 2009). The main damage came from the jaws, which fed on the immobilized prey.
Pack hunting: myth?
The idea that dromaeosaurids hunted in coordinated packs like wolves, popularized by the film Jurassic Park (1993), was challenged by Roach & Brinkman (2007). Analyzing sites like the Antlers Quarry in Oklahoma, where multiple Deinonychus specimens were found alongside a Tenontosaurus, they found evidence of cannibalism: Deinonychus teeth embedded in bones of other Deinonychus. This suggests behavior more similar to Komodo dragons: opportunistic group feeding with aggression between individuals, without real cooperation. Komodo dragons congregate around large carcasses and frequently attack each other. Studies of dental isotopes and tooth wear patterns at multi-individual theropod sites support this interpretation (Frederickson et al., 2020).
The hunter vs. scavenger debate
Jack Horner argued for years that T. rex was primarily a scavenger, based on its atrophied arms, powerful sense of smell (useful for finding carcasses), and relatively low speed. Most paleontologists today reject this hypothesis as exclusive: no known large terrestrial predator is an obligate scavenger (not even vultures are exclusively scavengers). The current consensus is that T. rex was an opportunistic predator, like lions and hyenas, hunting when possible and feeding on carcasses when available. The healed tooth in the hadrosaur is the strongest evidence of active predation.



How long did they live?
Paleontologists determine dinosaur age through bone histology: cutting long bones into thin sections (30-100 micrometers) and counting growth rings under a microscope, like in trees. Each ring represents approximately one year of life.
How bone histology works
The method consists of extracting a cross-section from a long bone (usually the tibia or femur), embedding it in resin, cutting it into ~30-micrometer slices with a microtome, and analyzing it under polarized light. Lines of Arrested Growth (LAGs) appear as dark, dense rings formed during periods of reduced growth (winter, dry season). Between LAGs, the bone tissue can be fibrolamellar (rapid growth) or lamellar (slow growth). The thickness between consecutive LAGs decreases as the animal ages, forming a sigmoidal growth curve: slow growth in infancy, explosive acceleration in adolescence, deceleration at maturity, and stagnation in old age. One of the method's problems is that bone remodeling (reabsorption and redeposition of bone tissue by the body) erases internal LAGs over the course of life, underestimating the age of older animals. Sauropods are particularly affected by this.
Theropod growth curves
The specimen "Sue" of T. rex died at approximately 28 years old and reached adult size around age 19. At peak growth, between ages 14 and 18, it gained about 2.1 kg per day (Erickson et al., 2004), the equivalent of going from ~500 kg to over 5,000 kg in just 4 years. This explosive growth is one of the strongest pieces of evidence for elevated metabolism. In comparison, Albertosaurus reached adult size at ~16 years and lived to ~28, while Gorgosaurus grew faster but died younger. The short lifespan of tyrannosaurids suggests they were high-metabolism, high-risk animals: growing fast, reproducing early, and dying relatively young. Psittacosaurus, a basal ceratopsian of 2 meters, is the non-avian dinosaur with the most histological data available. Erickson et al. (2009) analyzed dozens of specimens and determined it reached maturity at 8-9 years and lived to ~10-11 years.
Early reproduction
Sexual maturity came before skeletal maturity. Medullary bone, a calcium-rich reproductive tissue that today exists only in female birds during the egg-laying period (providing calcium for eggshells), was found in T. rex, Allosaurus, and Tenontosaurus, all in specimens still actively growing (Lee & Werning, 2008). This means dinosaurs were already reproducing before they stopped growing: a pattern closer to birds than to reptiles, which typically reach sexual maturity only after complete growth. Early reproduction is a classic strategy of animals with high mortality: starting to reproduce early increases the chances of leaving descendants before dying.
Juvenile mortality and life table
The study by Woodward et al. (2015) with 50 Maiasaura tibiae is the most complete example of paleodemography in dinosaurs. By determining the age of each individual through histology, the researchers constructed a complete life table, something normally only possible for living populations. The results revealed three distinct phases: 89.9% mortality in the first year of life, likely from predation, disease, and starvation; a 7-year "survival window" with relatively low mortality (12.7% per year), when the animals were already large enough to avoid most predators; and senescence after age 8, when the rate climbed to 44.4%, probably due to accumulated injuries, disease, and physiological wear. Maiasaura reached 2.3 tons in just 8 years.
Pathologies: what diseased bones reveal
Dinosaur bones frequently preserve signs of disease and injury. "Sue" suffered from gout, bone infections (osteomyelitis), and possible infected bites from other T. rex. Healed fractures in ribs and vertebrae are common in large theropods, indicating they survived injuries that would be fatal to smaller animals. In Allosaurus, over 50% of specimens from the Cleveland-Lloyd Quarry show healed fractures. These pathologies provide information about longevity (the animal lived long enough to heal), behavior (intraspecific fighting), and even diet (gout is associated with purine-rich diets, found in meat).
~28
years
T. rex
~8
years to adult
Maiasaura
30-70
years (estimated)
Sauropods


How intelligent were they?
Measuring the intelligence of extinct animals is one of paleontology's greatest challenges. The brain, made of soft tissue, does not preserve in fossils. What we have are endocasts: natural or digital casts (via CT scanning) of the cranial cavity, which reveal the general size and shape of the brain.
Encephalization Quotient (EQ)
The EQ, proposed by Harry Jerison in 1973, measures the ratio between brain mass and the expected mass for an animal of that body size. The logic is that larger animals need bigger brains simply to control their bodies: what "remains" beyond that minimum can be correlated with cognitive capabilities. Among non-avian dinosaurs, Troodon formosus holds the highest recorded EQ: its brain was about six times larger than predicted for a reptile of the same body size, a value comparable to modern emus and ostriches. Troodon had large forward-facing eyes (binocular vision), retractable toe claws, and prehensile hands with partial thumb opposition. At the opposite extreme, sauropods like Brachiosaurus carried brains of only 70 to 100 cm³ in bodies over 30 tons (for comparison, a domestic cat has ~25 cm³ in a 4 kg body). The EQ has serious limitations: it assumes the brain-body relationship follows a single formula for all animals, which is not true. Birds, for example, pack more neurons per gram of brain than mammals.
The T. rex brain
T. rex had a surprisingly large brain (~400 cm³), with well-developed olfactory bulbs and cerebral hemispheres. The olfactory lobes occupied about 40% of the brain volume, indicating exceptional smell. The cerebellum, responsible for motor coordination, was also proportionally large, consistent with a predator that needed precise head and body movements during attack. The optic lobes were moderate, suggesting that vision, while good, was not the dominant sense.
The Herculano-Houzel controversy
In 2023, Brazilian neuroscientist Suzana Herculano-Houzel published a study estimating that T. rex could have had ~3.3 billion neurons in the telencephalon, a number comparable to baboons and capuchin monkeys, animals capable of using tools and solving complex problems. If correct, this would imply that T. rex was cognitively sophisticated, possibly capable of behaviors previously considered exclusive to primates. The methodology, however, was severely criticized by Gutierrez-Ibañez et al. (2023) and by Caspar et al. (2024). The main criticisms: Herculano-Houzel used neuronal scaling equations derived from birds to estimate neurons in reptiles. Birds pack neurons far more densely than reptiles. Using the bird equation drastically overestimates the neuron count. The critics recalculated using reptile equations and arrived at ~1.5 billion neurons, a number comparable to large crocodiles, not primates. The brain size used by Herculano-Houzel was also questioned: she used the total endocast volume, but in reptiles the brain occupies only 30-50% of the cranial cavity (the rest is filled by meninges and fluid). This result remains highly controversial and most paleontologists take a cautious position.
Myth: the "second brain"
In the 19th century, O.C. Marsh observed that the neural canal in the sacral region of Stegosaurus was 20 times larger than the cranial cavity. The press of the era popularized the idea of a "second brain in the hip." In reality, this enlargement was not a second brain. Comparative studies with modern birds (Giffin, 1991) showed that the sacral enlargement in birds is occupied by the glycogen body, a glycogen-rich structure (energy reserve) whose exact function is still debated, but which is linked to the energy metabolism of the nervous system, not neural processing. Sauropods and ornithischians probably had analogous structures. The "second brain" is, therefore, a Victorian myth that persists in popular culture.
Sensory capabilities
Endocasts and CT scans reveal information about dinosaur senses. The inner ear preserved in skulls allows estimation of auditory frequency range: Parasaurolophus was sensitive to low-frequency sounds (consistent with crest vocalizations), while small theropods like Troodon heard higher frequencies, useful for locating small prey. The flocculus, a cerebellar structure visible in endocasts, is proportional to gaze agility: animals with large flocculi can maintain stable gaze while moving their heads rapidly. Maniraptoran theropods (dromaeosaurids, troodontids) had proportionally larger flocculi than any other dinosaur, suggesting visual agility comparable to birds of prey.
Complex behavior
There is no confirmed evidence of tool use in any non-avian dinosaur. Problem-solving and social learning remain speculative, inferred only indirectly from bone accumulations suggesting gregarious behavior and parental care. What we know is that maniraptoran theropods, especially troodontids, had brains, senses, and manual dexterity compatible with relatively sophisticated behaviors, possibly comparable to modern crows or parrots. However, extrapolating cognition from anatomy is inherently uncertain, and claims about dinosaurs being "as intelligent as primates" should be treated with skepticism until more robust evidence emerges.



References
Bates, K.T. & Falkingham, P.L. (2012). Estimating maximum bite performance in Tyrannosaurus rex using multi-body dynamics. Biology Letters, 8(4), 660-664.
Bell, P.R. et al. (2017). Tyrannosauroid integument reveals conflicting patterns of gigantism and feather evolution. Biology Letters, 13(6), 20170092.
Caspar, K.R. et al. (2024). How smart was T. rex? Testing claims of exceptional cognition in dinosaurs. The Anatomical Record, 307(4), 1223-1241.
Chen, P. et al. (1998). An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature, 391, 147-152.
Chin, K. et al. (1998). A king-sized theropod coprolite. Nature, 393, 680-682.
Dawson, R.R. et al. (2020). Eggshell geochemistry reveals ancestral metabolic thermoregulation in Dinosauria. Science Advances, 6(7), eaax9361.
Eagle, R.A. et al. (2011). Body temperatures of modern and extinct vertebrates from ¹³C-¹⁸O bond abundances in bioapatite. PNAS, 108(25), 10255-10260.
Eberth, D.A. & Getty, M.A. (2005). Ceratopsian bonebeds: occurrence, origins, and significance. In Currie, P.J. & Koppelhus, E.B. (eds.), Dinosaur Provincial Park, Indiana Univ. Press.
Erickson, G.M. et al. (2004). Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature, 430, 772-775.
Erickson, G.M. et al. (2009). A life table for Psittacosaurus lujiatunensis. Proc. Royal Society B, 276, 3085-3090.
Fowler, D.W. et al. (2011). The predatory ecology of Deinonychus and the origin of flapping in birds. PLOS ONE, 6(12), e28964.
Frederickson, J.A. et al. (2020). Ontogenetic dietary shifts in Deinonychus antirrhopus. Palaeogeography, Palaeoclimatology, Palaeoecology, 552, 109780.
Giffin, E.B. (1991). Endosacral enlargements in dinosaurs. Modern Geology, 16, 101-112.
Grady, J.M. et al. (2014). Evidence for mesothermy in dinosaurs. Science, 344(6189), 1268-1272.
Gutierrez-Ibañez, C. et al. (2023). How smart was T. rex? The Anatomical Record.
Horner, J.R. & Makela, R. (1979). Nest of juveniles provides evidence of family structure among dinosaurs. Nature, 282, 296-298.
Hutchinson, J.R. & Garcia, M. (2002). Tyrannosaurus was not a fast runner. Nature, 415, 1018-1021.
Ibrahim, N. et al. (2014, 2020). Semiaquatic adaptations in a giant predatory dinosaur. Science, 345, 1613-1616; Nature, 581, 67-70.
Jerison, H.J. (1973). Evolution of the Brain and Intelligence. Academic Press.
Lee, A.H. & Werning, S. (2008). Sexual maturity in growing dinosaurs does not fit reptilian growth models. PNAS, 105(2), 582-587.
Li, Q. et al. (2012). Reconstruction of Microraptor and the evolution of iridescent plumage. Science, 335(6073), 1215-1219.
Manning, P.L. et al. (2009). Biomechanics of dromaeosaurid dinosaur claws. Proc. Royal Society B, 276, 1071-1079.
Pol, D. et al. (2021). Earliest evidence of herd-living and age segregation amongst dinosaurs. Scientific Reports, 11, 20023.
Prum, R.O. (1999). Development and evolutionary origin of feathers. J. Experimental Zoology, 285(4), 291-306.
Rayfield, E.J. et al. (2001). Cranial design and function in a large theropod dinosaur. Nature, 409, 1033-1037.
Roach, B.T. & Brinkman, D.L. (2007). A reevaluation of cooperative pack hunting in Deinonychus. Bulletin Peabody Museum, 48(1), 103-138.
Ruben, J.A. et al. (1996). The metabolic status of some Late Cretaceous dinosaurs. Science, 273, 1204-1207.
Stevens, K.A. (2006). Binocular vision in theropod dinosaurs. J. Vertebrate Paleontology, 26(2), 321-330.
Varricchio, D.J. & Horner, J.R. (1993). Hadrosaurid and lambeosaurid bone beds from the Two Medicine Formation. Canadian Journal of Earth Sciences, 30(5), 997-1006.
Woodward, H.N. et al. (2015). Maiasaura, a model organism for extinct vertebrate population biology. Paleobiology, 41(4), 503-527.
Xu, X. et al. (2012). A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature, 484, 92-95.