Kronosaurus
Kronosaurus queenslandicus
"Kronos lizard from Queensland"
About this species
Kronosaurus queenslandicus was one of the largest pliosaurids of the Early Cretaceous, with an estimated length of 9 to 11 meters and a skull 2.2 meters long, one of the largest of any prehistoric reptile. It was not a dinosaur but a marine reptile of the clade Pliosauridae. It lived in the Eromanga Sea that covered inland Australia during the Aptian-Albian (~115-100 Ma). The nickname 'Plasterosaurus' reflects the controversial Harvard reconstruction (MCZ 1285), where eight extra plaster vertebrae were added to the specimen, inflating the length from 10.5 to 12.8 meters. With conical teeth up to 7 centimeters, it was the dominant apex predator of its environment, capable of attacking plesiosaurs, sea turtles, and large fish.
Geological formation & environment
Kronosaurus queenslandicus is found mainly in the Toolebuc Formation (late Aptian, ~100-95 Ma) and the Allaru Mudstone (Albian, ~100-95 Ma), both belonging to the Rolling Downs Group of Queensland. The Toolebuc Formation is famous for its calcareous nodules that preserve fish, cephalopods, and marine reptiles in excellent condition. The MCZ 1285 (Harvard) specimen was collected near Hughenden, Queensland, while specimen KK F0630 (mandible) came from the Julia Creek region. Both formations represent deposits of the Eromanga Sea, the epicontinental sea that covered inland Australia during the Early Cretaceous.
Image gallery













Life reconstruction of Kronosaurus queenslandicus by Nobu Tamura, showing the animal with four flippers and the enormous head in proportion to the body.
Nobu Tamura, CC BY-SA 3.0
Ecology and behavior
Habitat
Kronosaurus queenslandicus inhabited the Eromanga Sea, a shallow epicontinental sea (50-200 m) that flooded inland Australia during the Aptian-Albian (~115-100 Ma). The waters were warm and slightly brackish, with high biological productivity. The Toolebuc Formation and Allaru Mudstone, where the fossils were found, represent shallow to moderately deep sea deposits. The environment was tropical to subtropical, with surface water temperatures above 25°C and a rich fauna of ammonites, fish, turtles, and plesiosaurs.
Feeding
Kronosaurus was an apex carnivore, with conical teeth up to 7 centimeters capable of penetrating and holding large prey. The diet included plesiosaurs (evidenced by bite marks on fossil bones), sea turtles (the more rounded posterior teeth could crush shells), large fish, and cephalopods. The powerful mandible with long symphysis and premaxillary fang teeth indicate an ambush predator capable of capturing large prey. The skull size (20% of body length) maximized bite force.
Behavior and senses
Based on morphology and modern analogs, Kronosaurus was likely an ambush predator, using four flippers for rapid acceleration movements in attacks. There is no evidence of gregarious behavior. The hydrodynamic body shape suggests diving capability to seek prey at different depths. The closest modern ecological analogs are orcas and large sharks, with high-speed attack strategies on large prey.
Physiology and growth
Kronosaurus was likely mesothermic or endothermic, with a high metabolism needed to sustain the active large predator lifestyle. Bone histology of related pliosaurids indicates rapid growth and high metabolic rate. The large skull (2.2 m in length) generated one of the highest bite forces of any Mesozoic animal. The dentition with continuously replaceable teeth ensured maintenance of predatory capability throughout the animal's life.
Paleogeography
Continental configuration

Scotese & Vérard · CC BY 4.0 · Cretaceous, ~105 Ma
Fossil sites
Slate Weasel, CC BY 4.0
During the Aptiano-Albiano (~115–100 Ma), Kronosaurus queenslandicus inhabited Laramidia, the western half of present-day North America, separated from the east by the Western Interior Seaway, a shallow sea dividing the continent. The continents were in very different positions: India was drifting toward Asia, Antarctica was still connected to Australia, and South America was an isolated island.
Bone Inventory
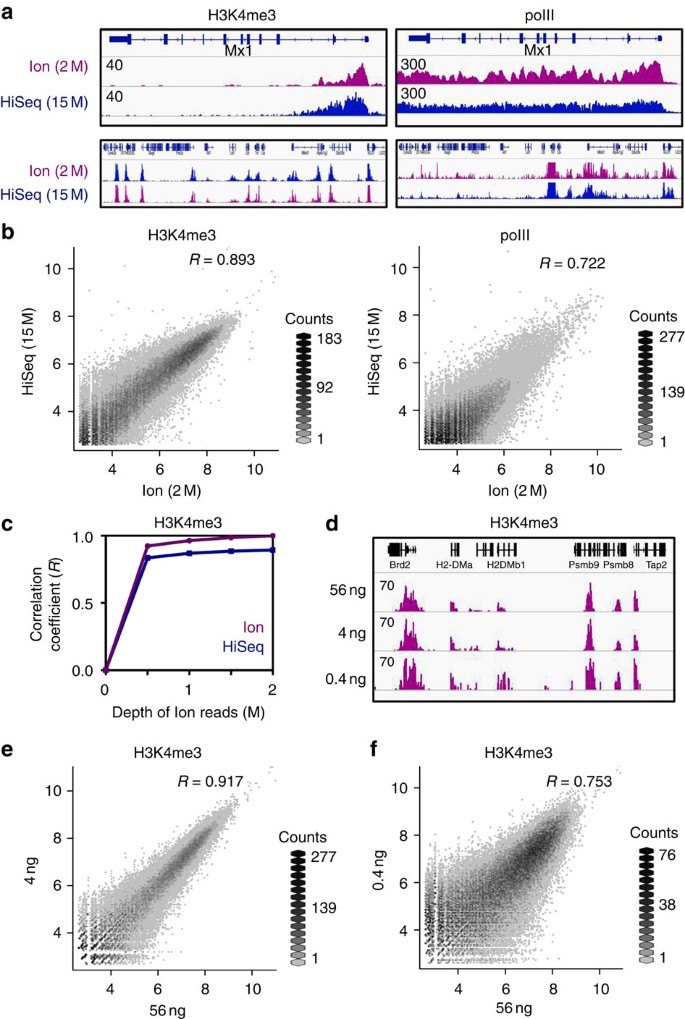
The MCZ 1285 specimen (Harvard) is the most complete, preserving a skull (2.2 m basal length), mandible (2.6 m), partial vertebral column, ribs, and flippers. However, 8 vertebrae were reconstructed in plaster by Romer and Lewis in 1959, making length estimates uncertain. Specimen QM F18827 (Queensland Museum), proposed as the neotype, preserves the skull in dorsal view. The 65% completeness reflects the composite of all known specimens.
Found elements
Inferred elements
Scientific Literature
15 papers in chronological order — from the original description to recent research.
A new gigantic marine reptile from the Queensland Cretaceous, Kronosaurus queenslandicus, new genus and species
Longman, H.A. · Memoirs of the Queensland Museum
Original description of Kronosaurus queenslandicus by Heber Albert Longman, based on a mandibular symphysis fragment found near Hughenden, Queensland. Longman recognizes the fossil as a giant pliosaurid and names the new genus and species. The work is fundamental as the taxonomic starting point, although the holotype material is fragmentary. Longman estimated the animal as one of the largest marine reptiles that ever lived, an estimate that has withstood the test of time.
On the skull of Kronosaurus queenslandicus Longman
White, T.E. · Occasional Papers of the Boston Society of Natural History
First detailed description of the Kronosaurus queenslandicus skull based on specimen MCZ 1285 collected by the Harvard expedition to Queensland in 1931. White documents cranial anatomy in detail, describing the skull roof bones, orbital orbits, nostrils, and tooth characteristics. This work established the basic knowledge of Kronosaurus cranial morphology that persisted for decades, until revised by McHenry and other researchers.
A mounted skeleton of the giant plesiosaur Kronosaurus
Romer, A.S. & Lewis, A.D. · Breviora
Romer and Lewis describe the mounting of the MCZ 1285 skeleton of Kronosaurus queenslandicus at the Harvard Museum of Comparative Zoology. The work documents the reconstruction process, including the controversial addition of about eight plaster vertebrae to complete the vertebral column. This decision artificially inflated the total animal length to 12.8 meters, a value that would only be corrected decades later by McHenry. The mounted specimen became one of the most famous paleontological displays in the United States.
Devourer of gods: the palaeoecology of the Cretaceous pliosaur Kronosaurus queenslandicus
McHenry, C.R. · PhD Thesis, University of Newcastle
Colin McHenry's doctoral dissertation providing the first comprehensive modern re-examination of Kronosaurus queenslandicus anatomy, taxonomy, and paleoecology. McHenry demonstrates that the Harvard MCZ 1285 specimen was reconstructed with eight extra plaster vertebrae, making the animal artificially ~3 meters longer. The work revises the skull (basal skull length = 2.2 m, mandible = 2.6 m), establishes that the actual length was ~10.5 m, and proposes QM F18827 as the neotype. It became the definitive anatomical reference for the species.

Marine reptiles from the Lower Cretaceous of South Australia: elements of a diverse fauna from the Eromanga Seaway
Kear, B.P. · Special Papers in Palaeontology
Kear documents the diverse marine reptile fauna from the Lower Cretaceous of South Australia, elements of a faunal assemblage from the Eromanga Seaway. The work includes pliosaurids related to Kronosaurus and provides paleoenvironmental context for the group. Kear describes the Eromanga Sea environment as a shallow epicontinental sea with warm waters and high diversity of potential prey for Kronosaurus. The associated fauna included elasmosaurid plesiosaurs, polycotylids, sea turtles, and large fish.
An archaic crested plesiosaur in opal from the Lower Cretaceous high-latitude deposits of Australia
Kear, B.P., Schroeder, N.I. & Lee, M.S.Y. · Biology Letters
Kear, Schroeder, and Lee describe a new opalized plesiosaur from the Lower Cretaceous of Australia, contemporaneous with Kronosaurus. The work documents the paleobiogeographic context of Australian marine reptiles from the Aptian-Albian and demonstrates the taxonomic diversity of the Eromanga Sea. This is relevant for Kronosaurus because it establishes the faunal context and indicates that opalized plesiosaurs were potential prey for the large pliosaurid.

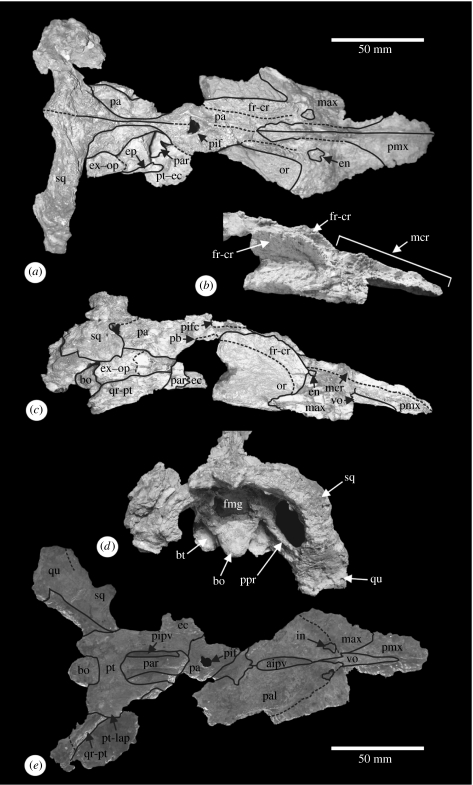
A new aeolodon (Pliosauridae) from the Jurassic of England and the phylogenetic relationships of thalassophonean pliosaurids
Foffa, D., Young, M.T., Brusatte, S.L., Graham, M.R. & Steel, L. · PeerJ
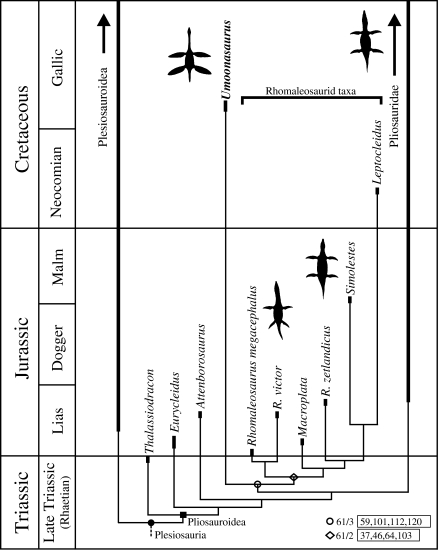
Foffa and colleagues describe a new pliosaurid (Aeolodon) from the Jurassic of England and conduct a phylogenetic analysis of thalassophoneans, including Kronosaurus queenslandicus. The analysis places Kronosaurus in a derived position within Brachaucheninae, within Thalassophonea. The work is relevant for understanding the evolutionary relationships of Kronosaurus within the family Pliosauridae and its position as one of the most derived thalassophoneans of the Early Cretaceous.

The mandible of Kronosaurus queenslandicus Longman, 1924 (Pliosauridae, Brachaucheninae), from the Lower Cretaceous of Northwest Queensland, Australia
Holland, T. · Journal of Vertebrate Paleontology
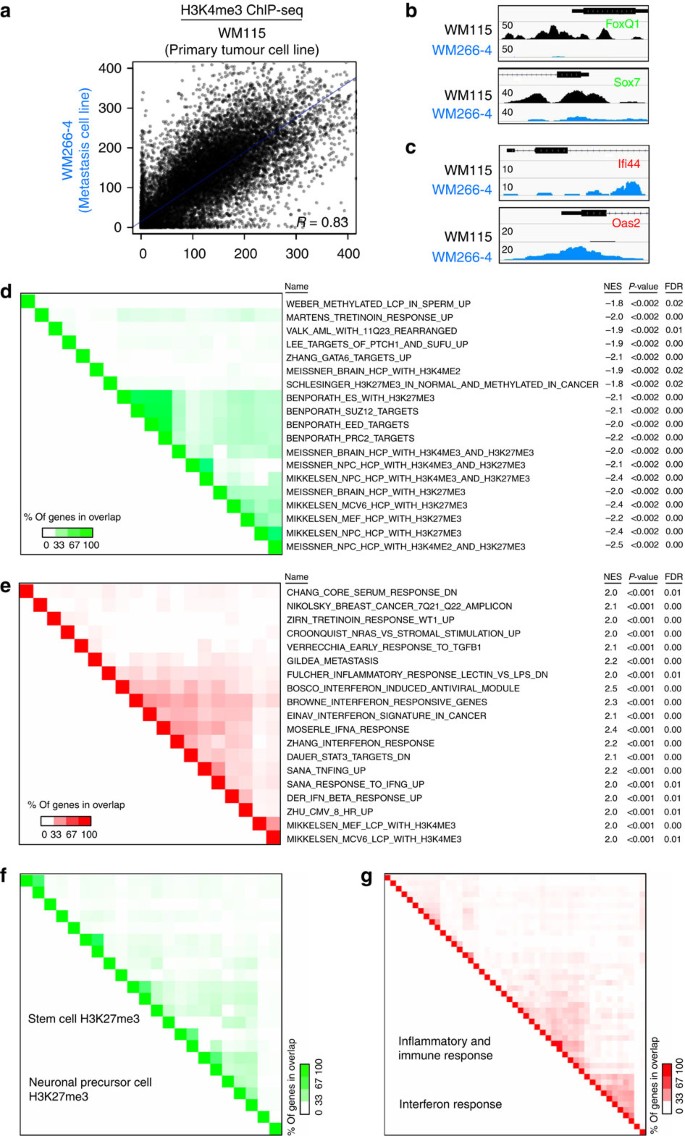
Holland presents the first complete description of the Kronosaurus queenslandicus mandible from specimen KK F0630, from the Allaru Mudstone of northwest Queensland. The work documents previously undescribed features, including a mandibular symphysis with lateral embayments to accommodate overhanging premaxillary teeth and a post-symphyseal constriction with embayments to accommodate upper maxillary teeth. Specimen KK F0630 significantly expands knowledge of the mandibular morphology of this species.

Giant pliosaurids (Sauropterygia; Plesiosauria) from the Lower Cretaceous peri-Gondwanan seas of Colombia and Australia
Poropat, S.F., Bell, P.R., Hart, L.J., Salisbury, S.W. & Kear, B.P. · Journal of Vertebrate Paleontology
Poropat and colleagues perform a comprehensive phylogenetic analysis of giant Lower Cretaceous pliosaurids from peri-Gondwanan Colombia and Australia, including Kronosaurus queenslandicus and K. boyacensis. The work formally proposes QM F18827 as the neotype of K. queenslandicus, resolving the taxonomic uncertainty around the fragmentary holotype. The analysis confirms Kronosaurus's position in Brachaucheninae and provides new data on the biogeography of giant Lower Cretaceous pliosaurids.


Endocranial anatomy of plesiosaurs (Reptilia: Sauropterygia) and its relevance for diving and hearing capabilities
Paulina-Carabajal, A., Cárdenas, G.H. & Reuil, S. · PeerJ
Paulina-Carabajal and colleagues analyze via CT scan the endocranial anatomy of plesiosaurs, including pliosaurids related to Kronosaurus. The work provides data on the sensory and diving capabilities of these marine reptiles. Endocranial analysis reveals that pliosaurids had relatively large olfactory lobes, suggesting acute smell for locating prey. Semicircular canals indicate balancing ability suitable for three-dimensional swimming.


Triassic-Early Cretaceous elasmosaurid plesiosaurs from Australia: new insights into their systematics and evolution
Kear, B.P. · Geological Journal
Kear reviews the systematics of Australian plesiosaurs from the Triassic to the Early Cretaceous, documenting the paleoenvironmental context for Kronosaurus and its contemporaneous fauna. The work provides data on the biogeography and evolution of marine reptiles on the Australian continental shelf during the Mesozoic. The review contextualizes Kronosaurus within the broader evolutionary history of Australian marine reptiles, demonstrating that the group had a continuous history on the continent since the Triassic.

Plasticity and Convergence in the Evolution of Short-Necked Plesiosaurs
Fischer, V., Benson, R.B.J., Zverkov, N.G., Soul, L.C., Arkhangelsky, M.S., Lambert, O., Stenshin, I.M., Uspensky, G.N. & Druckenmiller, P.S. · Current Biology
Fischer and colleagues perform a comprehensive analysis of the evolution of short-necked plesiosaurs (pliosaurids and polycotylids), demonstrating convergent evolution and placing Kronosaurus in a global phylogenetic context. The work is the most comprehensive on Pliosauridae evolution and reveals that the large-headed, short-necked body plan evolved independently in multiple lineages. The derived position of Kronosaurus within Brachaucheninae is confirmed by this analysis.


European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia
Neenan, J.M., Klein, N. & Scheyer, T.M. · Nature Communications
Neenan, Klein, and Scheyer phylogenetically analyze Sauropterygia, demonstrating the deep evolutionary roots of marine reptile groups. The work has implications for understanding Pliosauridae including Kronosaurus, by establishing the position of Pliosauridae within the broader phylogeny of Sauropterygia. The analysis demonstrates that the lineage leading to Kronosaurus diverged early in the evolutionary history of Sauropterygia and developed independent adaptations for large-scale predation.


Dental ontogeny and replacement in Pliosauridae
Sassoon, J., Foffa, D. & Marek, R. · Royal Society Open Science
Sassoon, Foffa, and Marek analyze via CT scan the dental ontogeny and replacement patterns in pliosaurids. The work has implications for growth rates and feeding strategies in Kronosaurus. The analysis demonstrates that pliosaurid teeth were continuously replaced throughout life, with new teeth erupting beneath existing ones. In Kronosaurus, the conical teeth up to 7 cm long were probably replaced at rates similar to those demonstrated in this study.


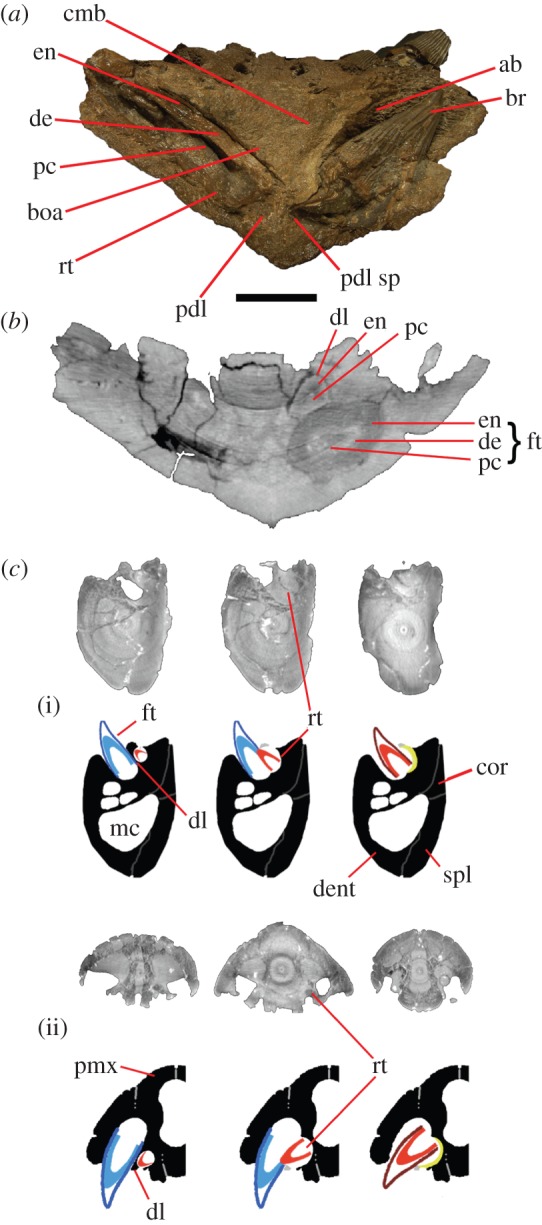
![Graph of alveolar size expressed as area (mm 2 ) in ( a ) Thalassiodracon (NHMUK R2039), ( b ) Peloneustes (NHM R8574), ( c ) P. kevani , ( d ) P. carpenteri and ( e ) P. westburyensis . Blue line, upper jaw; red line, lower jaw; yellow squares, alveoli in missing mandibular symphysis, as inferred by Benson et al . [ 34 ].](/images/species/kronosaurus-queenslandicus/papers/60badfeb0d-12.jpg)

Primaeval Oceans: Evolution of Jurassic-Cretaceous marine environments in Australia
Kear, B.P., Hamilton-Bruce, R.J. & Chapman, S.D. · Journal of the Geological Society of Australia (Special Issue)
Kear, Hamilton-Bruce, and Chapman document the evolution of Australian marine environments from the Jurassic to the Cretaceous, including the detailed paleogeography of the Eromanga Sea where Kronosaurus lived. The work describes the environmental conditions of the Eromanga Sea (shallow, warm, brackish waters), the associated fauna, and sedimentation patterns. It is the primary reference for understanding the specific paleoenvironment of Kronosaurus queenslandicus and contextualizing its fossils within Australian geological history.


Famous museum specimens
MCZ 1285
Museum of Comparative Zoology, Harvard University, Cambridge, Estados Unidos
The most famous Kronosaurus queenslandicus specimen, collected at Army Downs, near Hughenden, Queensland, by the Harvard expedition in 1930-1931. Mounted by Romer and Lewis in 1959 at 12.8 meters long, but with ~8 artificial plaster vertebrae added. McHenry (2009) estimated the actual length at ~10.5 m. The skull has a 2.2 m basal length and the mandible is 2.6 m. Permanent display at the museum since 1959.
QM F18827 (neótipo proposto)
Queensland Museum, Brisbane, Austrália
Specimen from the Toolebuc Formation (late Aptian), Queensland, proposed as the neotype of K. queenslandicus by Poropat et al. (2021) because it better represents the cranial anatomy of the species than Longman's (1924) fragmentary holotype. McHenry (2009) published the first detailed skull reconstruction based on this specimen. The reconstructed skull provides essential data on Kronosaurus cranial morphology.
KK F0630
Kronosaurus Korner, Richmond, Queensland, Austrália
Nearly complete mandible from the Allaru Mudstone (Albian), northwest Queensland, described by Holland (2018). Preserves the mandibular symphysis with unique lateral embayments and the post-symphyseal constriction. This specimen significantly expanded knowledge of Kronosaurus mandible morphology and is the most informative specimen for the mandibular anatomy of the species.
In cinema and popular culture
Kronosaurus has earned a permanent place in popular culture as the 'great white shark of the Australian Cretaceous', a role its colossal skull and sharp teeth justify with authority. In film and documentary series, it appeared most frequently in productions about 'prehistoric wildlife', prominently in Chased by Sea Monsters (BBC, 2003), where Nigel Marven virtually faces the animal in Australian Cretaceous seas. The IMAX documentary Sea Monsters: A Prehistoric Adventure (2007) brought Kronosaurus to large-format screens, consolidating its image as an apex predator. The controversial Harvard reconstruction (MCZ 1285), at 12.8 meters with plaster vertebrae, itself became an object of cultural fascination, with the nickname 'Plasterosaurus' gaining independent life on social media. In Australia, the species is a symbol of paleontological pride, with Kronosaurus Korner in Richmond, Queensland, becoming a tourist destination dedicated exclusively to the animal. The fact that the species gave its name to an entire museum attests to the power of a 2.2-meter skull over the human imagination.
Classification
Discovery
Fun fact
The most famous Kronosaurus specimen, mounted at Harvard in 1959, has eight false plaster vertebrae added by researchers, inflating the animal's length from about 10.5 to 12.8 meters. The informal nickname 'Plasterosaurus' still circulates among paleontologists as a reminder of this historical mistake.
Last reviewed: April 24, 2026