Dakotaraptor
Dakotaraptor steini
"Dakota raider, of Stein"
About this species
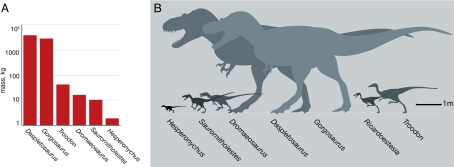
Dakotaraptor steini is a large dromaeosaurid from the Late Cretaceous (late Maastrichtian, about 66 to 67 million years ago) of the Hell Creek Formation, Harding County, South Dakota. Described by Robert DePalma, David Burnham, Larry Martin, Peter Larson and Robert Bakker in 2015, it reached 4.35 to 6 metres in length with an estimated body mass of 220 to 350 kilograms, making it the largest known Maastrichtian dromaeosaurid of North America and one of the largest of the family, alongside Utahraptor and Achillobator. The holotype PBMNH.P.10.113.T consists of a partial skeleton of a subadult to adult individual lacking a skull, discovered by Robert DePalma in 2005 in a Hell Creek fluvial channel no more than 20 metres below the Cretaceous-Palaeogene boundary. The ulna preserves about 15 ulnar papillae (quill knobs) of 8 to 10 millimetres in diameter, indicating attachment of large pennaceous feathers, and the second-toe 'sickle claws' typical of dromaeosaurids reach 24 centimetres along the outer curve. The tibia, 678 millimetres long, is the longest known in any dromaeosaurid, suggesting surprisingly gracile limb proportions for an animal of this size. In 2016, Arbour and colleagues demonstrated that the furculae originally described as part of the holotype were in fact entoplastra (parts of the shell) of the trionychid turtle Axestemys splendida, and DePalma et al. (2016) issued a corrigendum excluding those elements. The validity of the taxon remains debated: Cau (2023 and 2024) argued, in blog and phylogenetic analyses, that the remaining material may be a chimera combining ornithomimosaur, oviraptorosaur and therizinosaur elements, but this hypothesis has not yet been published in a peer-reviewed article.
Geological formation & environment
Hell Creek Formation, top of the unit, Upper Cretaceous (late Maastrichtian, ~66 to 67 Ma). The Hell Creek Formation crops out in Montana, North Dakota, South Dakota and Wyoming, recording fluvial and coastal-plain sediments deposited in the last 1 to 2 million years of the Cretaceous, immediately before the Cretaceous-Palaeogene (K-Pg) boundary dated at 66.043 ± 0.010 Ma by Sprain et al. (2018). The lithotype includes channel sandstones, floodplain mudstones, swamp coals and volcaniclastic bentonites. The unit overlies the Fox Hills Formation (marine) and underlies the Tullock / Fort Union Formation (Palaeocene). In Harding County, South Dakota, the Hell Creek Formation includes the Bull Creek sandstone and the Heikkila coal, and it is within this interval that the Dakotaraptor holotype was found, less than 20 metres below the K-Pg boundary. Associated fauna includes Tyrannosaurus rex, Triceratops horridus and prorsus, Torosaurus, Edmontosaurus annectens, Anzu wyliei, Pachycephalosaurus, Ankylosaurus magniventris, Thescelosaurus, Ornithomimus, Acheroraptor temertyorum (smaller dromaeosaurid) and troodontids such as Pectinodon, alongside crocodylians (Borealosuchus), turtles (including the trionychid Axestemys splendida, crucial for the Dakotaraptor 'furcula' controversy), mammals, squamates and birds.
Image gallery
















Life reconstruction of Dakotaraptor steini with full forearm plumage, inferred from the approximately 15 ulnar papillae (quill knobs) preserved on the holotype ulna.
Wikimedia Commons
Ecology and behavior
Habitat
Warm humid coastal plains of the North American interior during the late Maastrichtian, drained by meandering rivers crossing subtropical forests dominated by conifers (Taxodium, araucarians), palms (Arecaceae), cycadaceans, broad-leaved angiosperms (Dryophyllum) and pteridophytes. The coast of the epicontinental Western Interior Seaway still existed during Hell Creek deposition but was retreating rapidly. The unit includes active fluvial channels, hydromorphic floodplains, swamps and ephemeral ponds. The fauna includes Tyrannosaurus rex as dominant predator, Triceratops and Torosaurus among ceratopsids, Edmontosaurus annectens among hadrosaurids, Ankylosaurus and Denversaurus as ankylosaurs, Pachycephalosaurus and Thescelosaurus, and, among small theropods, Acheroraptor, Anzu, Ornithomimus and Troodon / Pectinodon. Dakotaraptor coexisted with all this fauna in the last hundreds of thousands of years of the Cretaceous.
Feeding
Active predator, strict carnivore. Its intermediate size between juvenile Tyrannosaurus rex and small theropods such as Acheroraptor suggests that Dakotaraptor occupied a mesopredator niche, possibly exploiting prey beyond the reach of juvenile tyrannosaurids but at an advantage over Acheroraptor and Troodon. Likely prey included Thescelosaurus, juvenile Pachycephalosaurus, ornithomimosaurs such as Ornithomimus and juvenile hadrosaurids. The 24 cm second-toe sickle claw and the gracile hindlimbs with a very long tibia suggest both active predation on open terrain and the capacity to grasp and subdue prey, as hypothesised by Fowler et al. (2011) for Deinonychus.
Behavior and senses
No tracks or sites are unequivocally attributable to Dakotaraptor, and the reduced post-corrigendum hypodigm limits direct inferences. By analogy with other eudromaeosaurs, solitary hunting or sporadic pair behaviour is possible, with rapid running on open terrain and attack with the sickle claw to subdue prey. The presence of ulnar papillae indicating forearm plumage suggests that visual display, sheltering over nests and thermoregulation were important roles of the feathers in the adult animal, independent of any flight capability, nonexistent given the body size.
Physiology and growth
Dakotaraptor was an exceptionally large dromaeosaurid with unusually gracile anatomy among the big representatives of the family. The 678 mm tibia is the longest known in Dromaeosauridae, indicating proportionally long hindlimbs and potentially greater cursorial capacity than Utahraptor (robust) and Achillobator (massive). The ulnar papillae attest to large forearm feathers, homologous to the secondaries of modern birds. Bone histology for the taxon has not been published in detail, which is particularly sensitive given the identification controversy, but Burnham and colleagues announced in post-2015 conference talks that the femur and ulna microanatomy is consistent with dromaeosaurids.
Paleogeography
Continental configuration

Scotese & Vérard · CC BY 4.0 · Cretaceous, ~105 Ma
During the Maastrichtiano tardio (~67–66 Ma), Dakotaraptor steini inhabited Laramidia, the western half of present-day North America, separated from the east by the Western Interior Seaway, a shallow sea dividing the continent. The continents were in very different positions: India was drifting toward Asia, Antarctica was still connected to Australia, and South America was an isolated island.
Bone Inventory
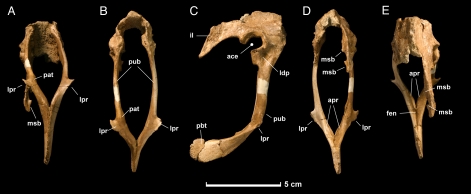
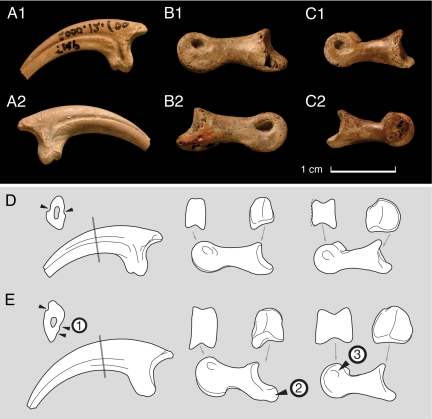
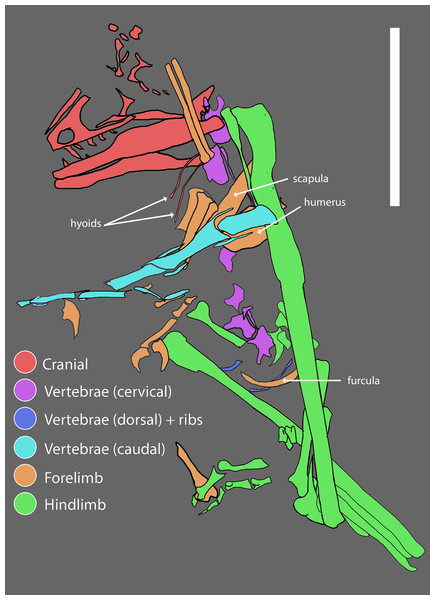
After the DePalma et al. (2016) corrigendum, holotype PBMNH.P.10.113.T consists basically of forelimbs, hindlimbs and caudal vertebrae of a single adult individual, without a skull or most of the axial column. The complete left tibia (678 mm) is the longest known in any dromaeosaurid. The ulna preserves about 15 ulnar papillae, the famous quill knobs indicating direct attachment of large pennaceous feathers on the caudal margin of the forearm, comparable to those described for Velociraptor by Turner and colleagues in 2007. One of the most discussed aspects of the taxon's status is the limited access to the type material: the Palm Beach Museum of Natural History is described by several authors as a private collection, constraining independent re-analysis, and Cau (2023 to 2024) argues, still at blog level, that the material may represent a chimera.
Found elements
Inferred elements
Scientific Literature
15 papers in chronological order — from the original description to recent research.
Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana
Ostrom, J.H. · Bulletin of the Peabody Museum of Natural History 30: 1 to 165
Foundational monograph for modern Dromaeosauridae. The description of Deinonychus antirrhopus, with its second-toe 'terrible claw' and bird-like manus, revived the hypothesis of dinosaurian ancestry for birds and is an indispensable basis for interpreting the anatomy of Dakotaraptor.


A large dromaeosaur (Theropoda) from the Lower Cretaceous of eastern Utah
Kirkland, J.I., Burge, D. e Gaston, R. · Hunteria 2(10): 1 to 16
Original description of Utahraptor ostrommaysorum, the giant dromaeosaurid from the Lower Cretaceous Cedar Mountain Formation of Utah. With approximately 7 metres of body length and skull bones 2.5 times larger than those of Deinonychus, it was for two decades the largest known dromaeosaurid, until equalled by Dakotaraptor in 2015.


A new maniraptoran theropod, Achillobator giganticus (Dromaeosauridae), from the Upper Cretaceous of Burkhant, Mongolia
Perle, A., Norell, M.A. e Clark, J.M. · Contributions of the Mongolian-American Paleontological Project
Description of Achillobator giganticus, a Mongolian Upper Cretaceous dromaeosaurid (~96 to 89 Ma), with a robust skeleton, primitive vertically oriented pubis and 4.5 to 5 metre body size. Key reference for understanding the diversity of large eudromaeosaurs and for comparing the ecology of a massive predator with the gracile build of Dakotaraptor.

Feather quill knobs in the dinosaur Velociraptor
Turner, A.H., Makovicky, P.J. e Norell, M.A. · Science 317(5845): 1721
First direct demonstration of quill knobs (ulnar papillae) in a nonavian dromaeosaurid, based on the ulna of Velociraptor mongoliensis (IGM 100/981). The feature is identical to that observed by DePalma et al. (2015) in Dakotaraptor, and is the basis for interpreting that giant Maastrichtian raptors had feathered forearms despite being far too large to fly.
A microraptorine (Dinosauria-Dromaeosauridae) from the Late Cretaceous of North America
Longrich, N.R. e Currie, P.J. · Proceedings of the National Academy of Sciences 106(13): 5002 to 5007
Description of Hesperonychus elizabethae, the smallest known nonavian dromaeosaurid from North America, from the Dinosaur Park Formation (Campanian, Alberta). Extends the temporal range of Microraptorinae by 45 million years and sets the context for discussing the body-size diversity of Late Cretaceous North American dromaeosaurids, of which Dakotaraptor represents the opposite extreme.
A review of dromaeosaurid systematics and paravian phylogeny
Turner, A.H., Makovicky, P.J. e Norell, M.A. · Bulletin of the American Museum of Natural History 371: 1 to 206
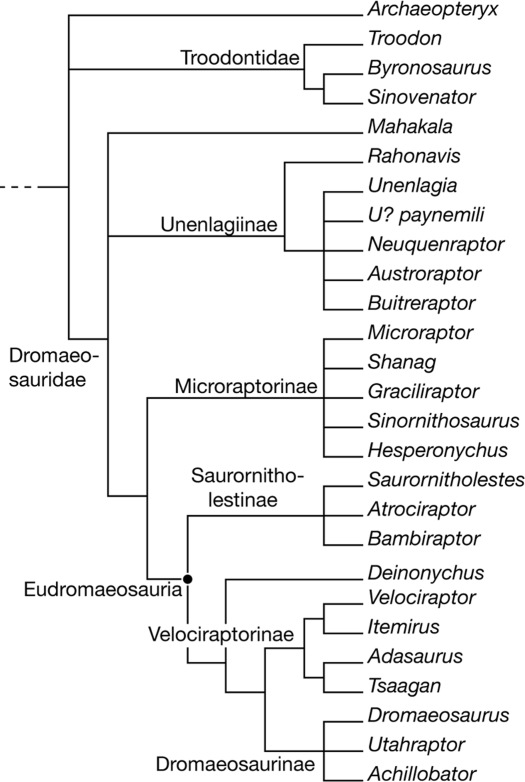
Monographic review of Dromaeosauridae systematics and paravian phylogeny. Defines Eudromaeosauria as the clade comprising Dromaeosaurinae, Velociraptorinae and Saurornitholestinae, the taxonomic framework within which Dakotaraptor was originally placed in 2015. Matrix foundation for later analyses by Jasinski (2020) and Currie and Evans (2019).
A new dromaeosaurid (Dinosauria: Theropoda) with Asian affinities from the latest Cretaceous of North America
Evans, D.C., Larson, D.W. e Currie, P.J. · Naturwissenschaften 100(11): 1041 to 1049
Description of Acheroraptor temertyorum based on cranial material from the Hell Creek Formation of Montana. Previously considered the only Hell Creek dromaeosaurid, it was joined two years later by Dakotaraptor, demonstrating that the formation hosted at least two contemporaneous dromaeosaurids in very different size classes.


The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation
DePalma, R.A., Burnham, D.A., Martin, L.D., Larson, P.L. e Bakker, R.T. · Paleontological Contributions 14: 1 to 16
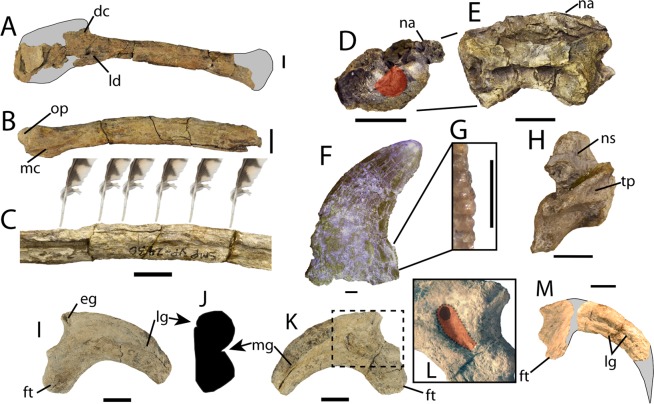
Original description of Dakotaraptor steini based on holotype PBMNH.P.10.113.T, collected by Robert DePalma in 2005 in the upper part of the Hell Creek Formation, Harding County, South Dakota, less than 20 metres below the Cretaceous-Palaeogene boundary. Key points: body size of about 5.5 metres and mass of about 350 kg, tibia of 678 mm (the longest known in Dromaeosauridae), about 15 ulnar papillae indicating large forearm feathers, and three elements described as furculae. Phylogenetic analysis placed Dakotaraptor as a eudromaeosaur close to Dromaeosaurus. This description is the primary reference and must be cited together with the 2016 corrigendum and the Arbour et al. (2016) critique.
The furculae of the dromaeosaurid dinosaur Dakotaraptor steini are trionychid turtle entoplastra
Arbour, V.M., Zanno, L.E., Larson, D.W., Evans, D.C. e Sues, H.-D. · PeerJ 4: e1691
Crucial taxonomic correction. The authors demonstrate that the three 'furculae' attributed to Dakotaraptor steini in the original paper, including the element incorporated into the holotype, are not theropod bones but rather entoplastra (central plates of the plastron) of the trionychid turtle Axestemys splendida, common in the Hell Creek Formation. The identification is based on the flattened V/U shape, the absence of a distinct epicleidial head, the spongy microanatomy lacking laminated pattern, and the perfect congruence with Axestemys known from the same site. The paper compelled the original authors to issue the 2016 corrigendum.

Corrigendum to: The first giant raptor (Theropoda: Dromaeosauridae) from the Hell Creek Formation
DePalma, R.A., Burnham, D.A., Martin, L.D., Larson, P.L. e Bakker, R.T. · Paleontological Contributions 16: 1 to 2
Formal corrigendum by the original authors, published 2 December 2016. Accepts the Arbour et al. (2016) reidentification of the 'furculae' as entoplastra of Axestemys splendida. Excludes from the Dakotaraptor hypodigm the furcula previously attributed to the holotype and the referred specimens KUVP 152429 (exclusively turtle material) and NCSM 13170. The reduced hypodigm remains the basis for all subsequent discussion of the taxon.
Cranial anatomy of new specimens of Saurornitholestes langstoni (Dinosauria, Theropoda, Dromaeosauridae) from the Dinosaur Park Formation (Campanian) of Alberta
Currie, P.J. e Evans, D.C. · The Anatomical Record 303(4): 691 to 715
Description of the cranial anatomy of a nearly complete Saurornitholestes langstoni skeleton from the Dinosaur Park Formation (Campanian, Alberta). Includes a new phylogenetic analysis in which Dakotaraptor is retained within Eudromaeosauria, but at a node sensitive to removal of unverifiable characters, in line with later critiques by Cau.


A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight
Hartman, S., Mortimer, M., Wahl, W.R., Lomax, D.R., Lippincott, J. e Lovelace, D.M. · PeerJ 7: e7247
Description of the troodontid Hesperornithoides miessleri and a large phylogenetic analysis of Paraves. In the Hartman and colleagues scheme, Dakotaraptor was recovered within Unenlagiidae (South American and Antarctic dromaeosaurids) rather than Eudromaeosauria. This topological instability is one of the main reasons why the position of Dakotaraptor remains contested.

New Dromaeosaurid Dinosaur (Theropoda, Dromaeosauridae) from New Mexico and biodiversity of dromaeosaurids at the end of the Cretaceous
Jasinski, S.E., Sullivan, R.M. e Dodson, P. · Scientific Reports 10: 5105
Description of Dineobellator notohesperus, the first diagnostic dromaeosaurid from the latest Cretaceous of the southern United States (New Mexico). The phylogenetic analysis recovers Eudromaeosauria with Saurornitholestinae, Dromaeosaurinae and Velociraptorinae, with Dakotaraptor and Acheroraptor among the Maastrichtian terminals. This is the most cited published cladogram for placing Dakotaraptor in recent trees.
Osteology and reassessment of Dineobellator notohesperus, a southern eudromaeosaur (Theropoda: Dromaeosauridae: Eudromaeosauria) from the latest Cretaceous of New Mexico
Jasinski, S.E. · The Anatomical Record 306(7): 1641 to 1708
Osteological monograph of Dineobellator notohesperus with a broad revision of Eudromaeosauria. Discusses the body-size diversity of Maastrichtian dromaeosaurids, placing Dineobellator as a medium-sized eudromaeosaur and Dakotaraptor as the size extreme of the same clade. Reinforces, with more characters, that Dakotaraptor belongs within Eudromaeosauria, while acknowledging the topological instability highlighted by Hartman et al. (2019) and Cau.
Calibration of chron C29r: new high-precision geochronologic and paleomagnetic constraints from the Hell Creek region, Montana
Sprain, C.J., Renne, P.R., Clemens, W.A. e Wilson, G.P. · GSA Bulletin 130(9-10): 1615 to 1644
High-precision geochronological calibration (U-Pb CA-TIMS on zircon and 40Ar/39Ar) of the Hell Creek Formation in Montana. Places the K-Pg boundary at 66.043 ± 0.010 Ma and constrains deposition of the upper Hell Creek to less than 1 million years. Essential context to assert that Dakotaraptor lived in the final 100 to 400 thousand years of the Cretaceous, less than 20 m below the K-Pg boundary at the holotype site.


Famous museum specimens
PBMNH.P.10.113.T (holótipo de Dakotaraptor steini)
Palm Beach Museum of Natural History, Flórida, EUA
Holotype of Dakotaraptor steini, collected in 2005 in a fluvial channel of the upper Hell Creek Formation, Harding County, South Dakota, less than 20 metres below the Cretaceous-Palaeogene boundary. The material was deposited in the Palm Beach Museum of Natural History (Florida), described by several authors as a private collection, which has drawn criticism regarding access for independent re-analysis. After the DePalma et al. (2016) corrigendum, the furcula incorporated into the holotype was removed from the hypodigm, as it had been reidentified as entoplastron of the turtle Axestemys splendida by Arbour et al. (2016).
PBMNH.P.10.115.T e PBMNH.P.10.118.T (referidos)
Palm Beach Museum of Natural History, Flórida, EUA
Specimens referred in the 2015 original paper that remained in the hypodigm after the 2016 corrigendum. They are fragments and isolated bones whose association with Dakotaraptor is based on size and morphology criteria compatible with the holotype. Even so, Cau's analysis (2023 to 2024) raises doubts about the homogeneity of the set.
KUVP 156045 (referido)
University of Kansas Museum of Natural History (KU Biodiversity Institute), Lawrence, Kansas, EUA
One of the specimens originally referred to the taxon and retained after the corrigendum. Important because it is housed in an accessible public collection (KU), which enables independent reassessment. Specimen KUVP 152429, also referred in 2015, was excluded by the 2016 corrigendum as consisting exclusively of turtle material.
Réplicas e montagens reconstruídas
Palm Beach Museum of Natural History (Flórida) e exposições itinerantes
Reconstructed skeletal replicas using holotype material and casts of Utahraptor and Achillobator have been shown at educational events. No permanent official skeletal mount of Dakotaraptor was installed at a major public museum up to 2026.
Classification
Discovery
Fun fact
The most famous part of Dakotaraptor may not be the 24 cm sickle claw or the ulnar papillae that show plumage on an animal far too large to fly: it is the 'furcula that wasn't a furcula'. In 2015, DePalma and colleagues described three U-shaped bones as Dakotaraptor furculae, one of them part of the holotype. In February 2016, Arbour, Zanno, Larson, Evans and Sues showed that these bones are actually entoplastra (central plates of the plastron) of the trionychid turtle Axestemys splendida, common in the Hell Creek Formation. The original authors published a formal corrigendum in December 2016 accepting the correction. The episode became a palaeontology case study on the importance of independent review and access to type material, and is an inseparable part of the taxon's scientific identity.
Last reviewed: April 24, 2026