Comparative anatomy
Teeth that tell stories
What does a fossil tooth reveal about how an animal hunted? In paleontology, enamel is one of the richest pieces of the puzzle. From shape, serration, thickness and curvature, paleontologists reconstruct diet and hunting strategy with surprising precision. This page compares nine predators and shows that teeth that look similar under the microscope can reveal completely different behaviors.
How to read a predator tooth
Before comparing species, it helps to understand three variables paleontologists use to classify carnivorous teeth. Almost every difference we will see below appears in some combination of these three.
1. Cross-sectional shape
A tooth is ziphodont when it is laterally flattened, blade-shaped, like a knife. It is conidont when its cross-section is round, cone-shaped, like a spike. Blades cut, cones puncture. The choice between the two shapes typically indicates whether the predator attacked large prey (blade, for slicing) or slippery prey (cone, for puncturing and holding).
2. Serrations (denticles)
Tiny teeth along the edges, like a bread knife. The finer and more regular they are, the more specialized for cutting flesh. Carcharodontosaurids have serrations so regular that the group's name means "shark-toothed lizard" (from Carcharodon, the genus of the great white shark). Spinosaurids, in contrast, have reduced or absent serrations, a clear sign of a different diet.
3. Robustness and root
A thin, curved tooth is efficient for cutting, but breaks when it hits bone. A thick tooth, with a deep root in the jaw, withstands extremely high impact bites. T-Rex took this pattern to the extreme: thickened (incrassate) teeth with a root almost the same size as the crown, capable of bites of more than 3.5 tons of force without breaking.
School 1
The bone crushers
When a predator is large enough to bite into prey vertebrae and process entire carcasses, knife-thin teeth are useless: they would shatter. The evolutionary solution was to thicken everything. Tyrannosaurids, and T-Rex in particular, are the most extreme case of this strategy in the entire history of dinosaurs.


What the tooth tells us about hunting
T-Rex teeth were incrassate: the base is almost as wide as the tooth is long, and the enamel is thicker than that of any other known theropod. Erickson and colleagues (1996) and later finite element studies showed that this geometry distributes the stress of a brutal bite without fracturing the tooth.
The bite force was estimated by Bates and Falkingham (2012) and Gignac and Erickson (2017) at between 3,500 and 6,400 kgf, the highest of any known terrestrial animal, living or extinct. A combination of thickened teeth and extreme force enabled what paleontologists call "puncture-and-pull": bite with full force, puncturing flesh and bone, and pull the head back to tear off chunks. Tooth marks of T-Rex on fossils of Triceratops and Edmontosaurus show exactly this pattern.
The strategy had a cost: incrassate teeth are inefficient for cutting compared to thin blades. T-Rex compensated for this by replacing teeth throughout life (as did other theropods), in cycles of about 2 years per tooth, according to D'Emic et al. (2019).
School 2
The flesh slicers
Carcharodontosaurids and allosaurids followed the opposite strategy from T-Rex. Instead of thickening the tooth to withstand bites into bone, they thinned it for a brutal specialization: cutting flesh into deep slices. This "school" includes the largest predators that ever lived, and dominated the top of the food chain from the middle Jurassic to the middle Cretaceous.
Carcharodontosaurus saharicus
"Shark-toothed lizard", Middle Cretaceous (~99 to 94 Ma), North Africa

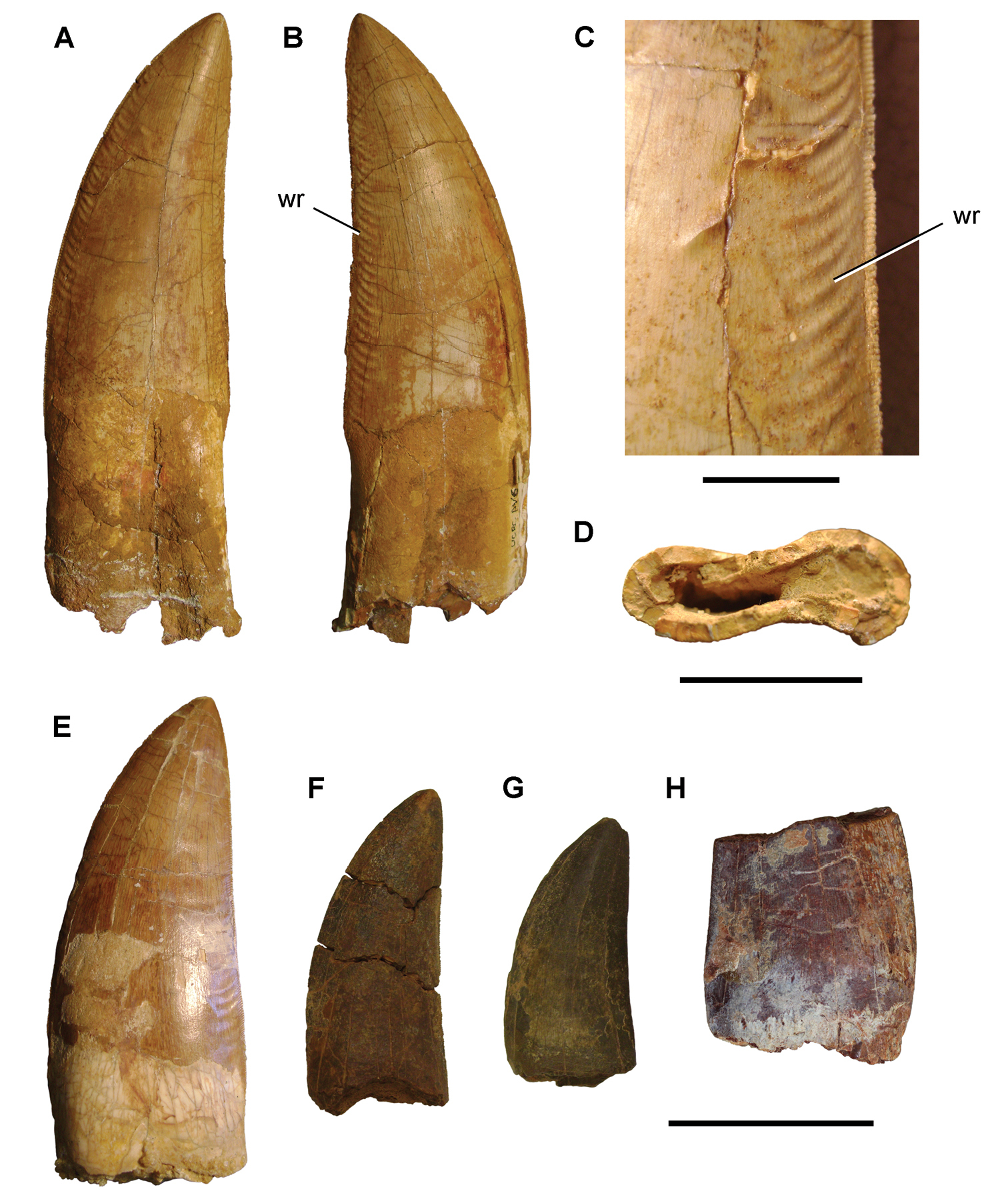
Giganotosaurus carolinii
Late Cretaceous (~99 to 95 Ma), Argentine Patagonia. South American cousin of Carcharodontosaurus.


Allosaurus fragilis
Late Jurassic (~155 to 145 Ma), North America. Older cousin of the carcharodontosaurids.

What the tooth tells us about hunting
For Carcharodontosaurus and Giganotosaurus, the interpretation is straightforward: teeth designed to slice flesh into deep cuts, especially of sauropods. Bite force studies on carcharodontosaurids (Bates and Falkingham, 2018) estimate something around 2,000 to 3,000 kgf, below T-Rex, but enough to penetrate the thick skin of titanosaurs. The strategy depended on the volume of blood lost by the prey: deep, repeated cuts until the prey collapsed, similar to behavior described today in great white sharks attacking seals.
Allosaurus follows the same general design, but with a lighter skull and proportionally smaller teeth. Rayfield (2001) proposed the famous "hatchet bite" hypothesis: the Allosaurus skull handled vertical loads well, but lateral loads poorly. The interpretation is that it used the skull like a hatchet, driving the head down with neck muscles onto the prey, instead of biting and pulling like T-Rex.
In all three cases, the serrated teeth left a characteristic pattern on bones: fine parallel marks, distinct from the pointed, broad marks left by T-Rex's incrassate teeth. This contrast is what allows paleontologists to identify the author of a bite on fossils more than 100 million years old.
School 3
The fish catchers
Spinosaurids completely break the pattern of carnivorous theropods. Instead of serrated blades, they have conical, smooth, unserrated teeth. The result looks like a crocodile tooth, and that is no coincidence: the function is the same, gripping slippery prey and holding it until swallowed.
Spinosaurus aegyptiacus
Middle Cretaceous (~99 to 93 Ma), North Africa. The largest known theropod, possibly semi-aquatic.


Baryonyx walkeri
Early Cretaceous (~125 Ma), England. Smaller and older cousin of Spinosaurus.

What the tooth tells us about hunting
Conical teeth without serrations are terrible for cutting flesh and excellent for puncturing and gripping. It is exactly the opposite of the slicer school. Baryonyx is the most conclusive case: the holotype described by Charig and Milner (1986) preserved partially digested scales and teeth of the fish Lepidotes in the animal's stomach, direct evidence of a piscivorous diet. It was the first dinosaur with fish stomach contents ever found.
For Spinosaurus, the evidence is more indirect but convergent. Ibrahim and colleagues (2014, 2020) reanalyzed fossils and proposed that Spinosaurus was the first known semi-aquatic theropod: paddle-shaped tail, high-positioned nostrils, dense bones for reduced buoyancy. The dental morphology fits this scenario. Beevor et al. (2021) studied the massive quantity of Spinosaurus teeth in fluvial Kem Kem deposits and concluded that the animal spent much of its life in water, feeding on fish like the prehistoric Onchopristis, a sawfish two meters long.
Summary of the school: the conical tooth is an ecological signature. A theropod with such teeth almost certainly ate fish or other slippery prey. The morphology was conserved in the spinosaurid lineage for more than 30 million years.
School 4
The small supporting actors
Not every predator relies primarily on its teeth. Dromaeosaurids, the group of Velociraptor and Deinonychus, have small, recurved, serrated teeth, similar to miniature versions of ziphodonts. But the main weapon was on the foot: the famous retractable sickle claw on the second toe. Teeth came afterward.


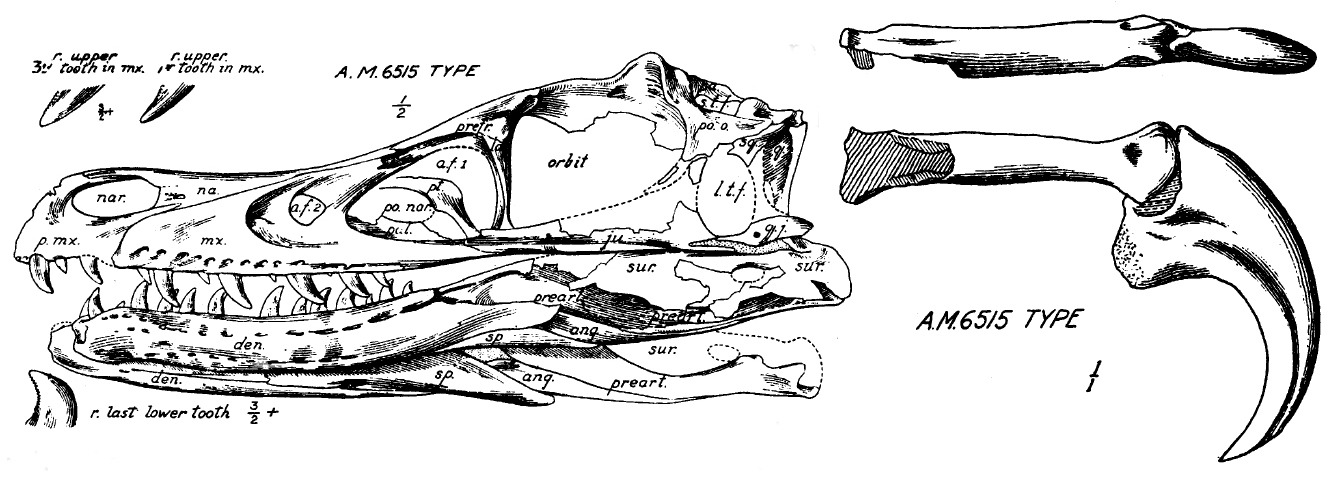
What the tooth tells us about hunting
Dromaeosaurids are the clearest example that teeth alone do not tell the whole story. Their main weapon was the retractable claw on the second toe of the foot, which could reach 13 cm in Deinonychus. Fowler et al. (2011) proposed the "raptor prey restraint" (RPR) hypothesis: the predator climbed onto smaller prey and used body weight plus claw to immobilize it, biting the live prey while it still struggled. This hypothesis is supported by behavioral studies of modern raptors (eagles, hawks), which kill their prey in exactly this way.
In this strategy, teeth function as processing tools, not hunting tools. They tear off chunks of flesh while the foot and weight hold the animal down. It is very different from T-Rex or Carcharodontosaurus, where the bite is the killing method itself.
The famous case of the "Fighting Dinosaurs", a Mongolian fossil discovered in 1971 with a Velociraptor entangled with a Protoceratops, shows the raptor's claw sunk into the herbivore's throat, and the teeth with no record of lethal use. The choreography of the hunt is all in the foot.
Outliers
Three cases that break the rule
Some species do not fit the main spectrum. They show that evolution is not linear: the same group (theropods) can produce an animal that eats fish, another that crushes bone, another that would hold prey like a felid, and even a theropod that turned herbivore.
Majungasaurus crenatissimus
Late Cretaceous (~70 to 66 Ma), Madagascar. The only large predator on the island.

Strategy: bite and hold
Snively et al. (2011) used finite element analysis to reconstruct the mechanics of the Majungasaurus skull. The conclusion: a strategy similar to that of modern felids, bite and hold the prey while neck musculature did the rest. The shorter, more robust teeth handled sustained loads during the struggle with the prey, not the single bite of the slicers.
The main prey was Rapetosaurus krausei, a titanosaur about 15 meters long. There are tooth marks of Majungasaurus on Rapetosaurus bones, identified by Rogers et al. (2007).
And there is a macabre detail: tooth marks of Majungasaurus on Majungasaurus bones, described by Rogers, Krause and Curry Rogers (2003) in Nature. It is the only well-documented case of cannibalism in non-avian dinosaurs. Was it active predation or carcass consumption? Still under debate.
Therizinosaurus, Erlikosaurus, Falcarius
Cretaceous (~125 to 70 Ma), Asia and North America. Theropods that turned herbivorous.
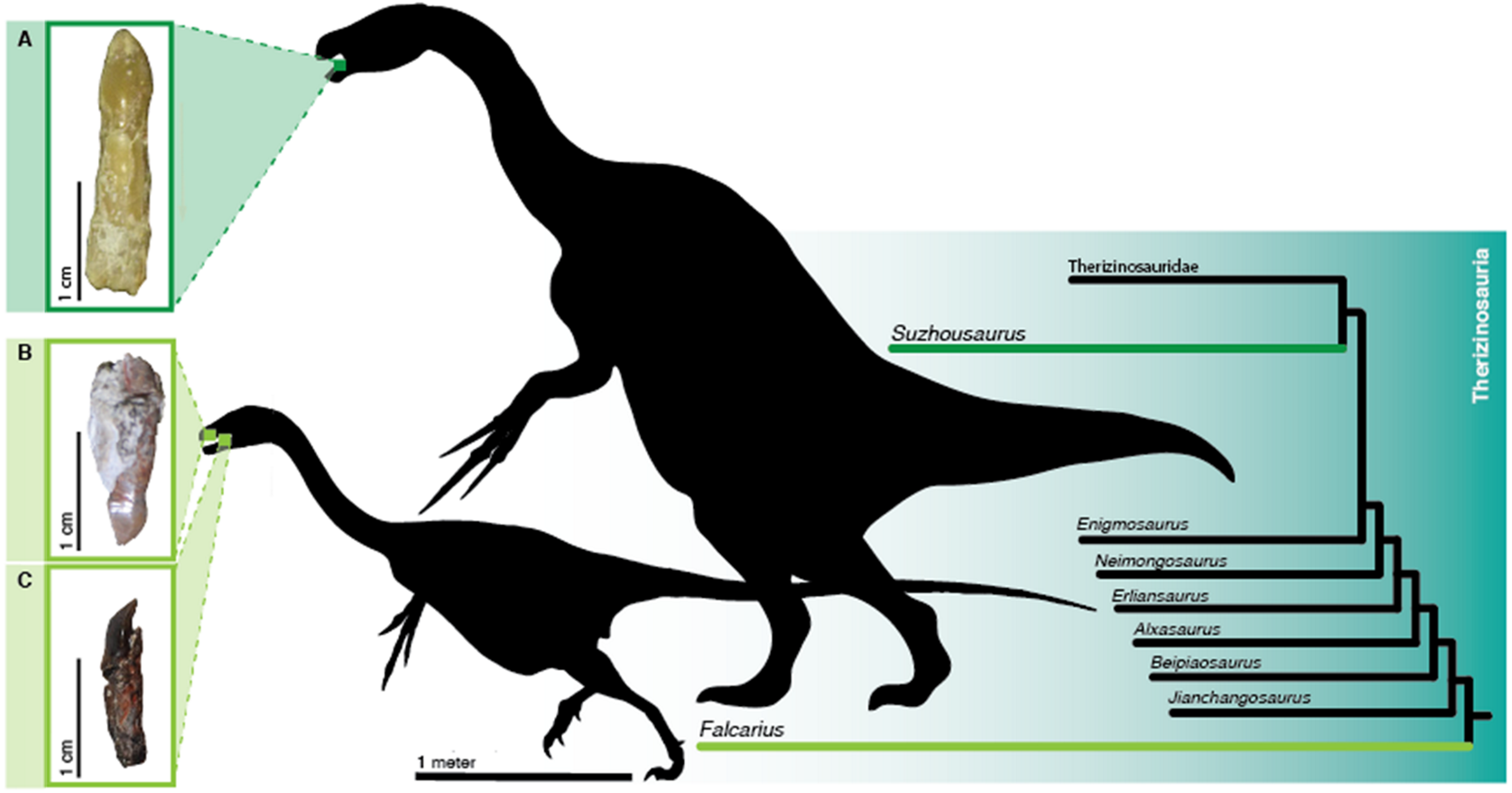

The evolutionary twist
Therizinosaurids were theropods, the same group as T-Rex and Velociraptor. Phylogenetically, they are close relatives of birds and dromaeosaurids. But during the Cretaceous, some lineage made the most radical possible transition: stopped being carnivorous and became herbivorous.
The evidence is in the teeth. Button et al. (2017) analyzed the histology of teeth from Falcarius and Suzhousaurus and showed that there were changes in the increment pattern of tissues compatible with a plant diet. Leaf-shaped teeth are ineffective for cutting flesh, but efficient at shredding vegetation. Combined with anterior keratinous beaks (as in birds), long necks and enormous bellies for gut fermentation, they make up a "plant-eating machine" disguised as a carnivore.
The case breaks a simple intuition: theropod is not synonymous with carnivore. The lineage is defined by general anatomy (bipedalism, hollow bones, etc.), not by diet. And the tooth, once again, is the signal of the change.
Carnotaurus sastrei
Late Cretaceous (~72 to 70 Ma), Argentine Patagonia. Short-faced abelisaurid with horns.

Strategy: fast bite with a powerful neck
Mazzetta and colleagues (1998, 2004, 2009) modeled the mechanics of the Carnotaurus skull and reached a curious result: it did not have a particularly strong bite. The estimated force is lower than that of an Allosaurus of similar size. But the neck was enormously muscular: the cervical vertebrae have muscle insertion areas comparable to those of bulls and bison.
The consensus interpretation: Carnotaurus delivered short, fast bites while moving, relying on the neck to generate impact force. The small teeth were sufficient for that kind of hunting, and the short face reduced the moment (lever arm) when striking. It brings to mind a lion running sideways after a gazelle and delivering a series of bites, instead of a T-Rex stopping prey with brute force.
Carnotaurus also had remarkably short, atrophied arms, even more so than T-Rex. The neck musculature and speed made up for everything. It is another reminder that teeth are only one piece: to understand how a predator hunted, you have to look at the entire skull, the neck, and the rest of the body.
Overview: five schools, nine predators
Each predator adapted its teeth to the type of prey, the size of its own body and what the rest of the anatomy offered. Small differences in geometry correspond to radically distinct hunting strategies.
| Predator | Tooth type | Hunting strategy | Typical prey |
|---|---|---|---|
| T-Rex | Banana, incrassate, deep root | Crushes bone (puncture-and-pull) | Hadrosaurs, Triceratops |
| Carcharodontosaurus | Very flat blade, serrated | Slices flesh into deep cuts | Sauropods (titanosaurs) |
| Giganotosaurus | Blade, identical to Carcharodontosaurus | Slices flesh, possibly in groups | Patagonian sauropods |
| Allosaurus | Blade, shorter and more robust | Hatchet bite (vertical strike) | Jurassic sauropods, stegosaurs |
| Spinosaurus | Conical, smooth, unserrated | River fishing (semi-aquatic) | Fish, sawfish (Onchopristis) |
| Baryonyx | Conical, long snout (gharial-like) | River fishing (confirmed by stomach) | Fish (Lepidotes) |
| Velociraptor / Deinonychus | Small, recurved, serrated | Sickle claw immobilizes, tooth processes | Protoceratops, small prey |
| Majungasaurus | Short, robust, rounded base | Bite and hold (felid-style) | Rapetosaurus, cannibalism |
| Therizinosaurus | Leaf-shaped, no sharp serration | Does not hunt: it is herbivorous | Leaves, fruits, possibly insects |
| Carnotaurus | Small for the animal's size | Fast bite with powerful neck | Medium prey, attacks on the move |
Conclusion
A fossil tooth is a document. From shape, serration, thickness and curvature, paleontologists reconstruct what the animal ate, how it attacked, and even social behaviors. The diversity of dental solutions among predatory dinosaurs is proof that the niche of "apex predator" was filled by very different strategies throughout the Mesozoic, from the American bone-crusher to the African river fisher, from the Madagascan island cannibal to the theropod that became herbivorous.
Sources
- Bates, K. T. & Falkingham, P. L. (2012). Estimating maximum bite performance in Tyrannosaurus rex. Biology Letters, 8(4), 660-664.
- Bates, K. T. & Falkingham, P. L. (2018). The importance of muscle architecture in biomechanical reconstructions of extinct animals. J. Anatomy.
- Beevor, T. et al. (2021). Taphonomic evidence supports an aquatic lifestyle for Spinosaurus. Cretaceous Research, 117, 104627.
- Brownstein, C. D. (2019). New records of theropods from the latest Cretaceous of New Jersey. Royal Society Open Science.
- Button, K., You, H., Kirkland, J. I. & Zanno, L. E. (2017). Incremental growth of therizinosaurian dental tissues. PeerJ, 5:e4129.
- Charig, A. J. & Milner, A. C. (1986). Baryonyx, a remarkable new theropod dinosaur. Nature, 324, 359-361.
- Cuff, A. R. & Rayfield, E. J. (2013). Feeding mechanics in spinosaurid theropods and extant crocodilians. PLOS ONE, 8(5).
- D'Emic, M. D. et al. (2019). Evolution of high tooth replacement rates in theropod dinosaurs. PLOS ONE, 14(11).
- Erickson, G. M. et al. (1996). Bite-force estimation for Tyrannosaurus rex from tooth-marked bones. Nature, 382, 706-708.
- Fowler, D. W. et al. (2011). The predatory ecology of Deinonychus and the origin of flapping in birds. PLOS ONE, 6(12).
- Gignac, P. M. & Erickson, G. M. (2017). The biomechanics behind extreme osteophagy in Tyrannosaurus rex. Scientific Reports, 7, 2012.
- Ibrahim, N. et al. (2014). Semiaquatic adaptations in a giant predatory dinosaur. Science, 345, 1613-1616.
- Ibrahim, N. et al. (2020). Tail-propelled aquatic locomotion in a theropod dinosaur. Nature, 581, 67-70.
- Mazzetta, G. V., Cisilino, A. P., Blanco, R. E. & Calvo, N. (2009). Cranial mechanics and functional interpretation of the horned carnivorous dinosaur Carnotaurus sastrei. Journal of Vertebrate Paleontology, 29(3), 822-830.
- Rayfield, E. J. et al. (2001). Cranial design and function in a large theropod dinosaur. Nature, 409, 1033-1037.
- Rogers, R. R., Krause, D. W. & Curry Rogers, K. (2003). Cannibalism in the Madagascan dinosaur Majungatholus atopus. Nature, 422, 515-518.
- Rogers, R. R. et al. (2007). Paleoenvironment and taphonomy of the Anembalemba Member, Maevarano Formation. SVP Memoir 8.
- Snively, E. et al. (2011). Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods. PeerJ.