Velociraptor
Velociraptor mongoliensis
"Ladrão veloz da Mongólia"
Sobre esta espécie
O Velociraptor mongoliensis era um dinossauro dromeossaurídeo de pequeno porte, com cerca de dois metros de comprimento e menos de 20 kg, que viveu no Campaniano tardio do Cretáceo, há 75 a 71 milhões de anos, na Mongólia. Ao contrário da representação cinematográfica em Jurassic Park, era aproximadamente do tamanho de um peru. Possuía penas confirmadas por evidências diretas de knobs de cálamos no rádio, uma garra falciforme elevada no segundo dedo do pé, e era provável caçador ágil de pequenos vertebrados no ambiente árido de dunas de areia da Formação Djadochta.
Formação geológica e ambiente
A Formação Djadochta (às vezes transliterada como Djadokhta) é uma unidade geológica do Campaniano tardio (~75-71 Ma) localizada no sul da Mongólia e na Mongólia Interior, China. Representa um ambiente desértico eólico com dunas de areia e correntes fluviais intermitentes, semiárido e quente. É uma das formações mais ricas em fósseis do Cretáceo tardio do mundo, preservando Velociraptor mongoliensis, Protoceratops andrewsi, Oviraptor philoceratops, Pinacosaurus grangeri e inúmeros mamíferos e lagartos. O famoso sítio Bayn Dzak ('Penhascos Flamejantes') é o local de descoberta do holótipo de Velociraptor (1923) e de Protoceratops (1922). A sedimentação eólica rápida, como deslizamentos de dunas, é responsável pela preservação excepcional de espécimes como os Dinossauros Lutadores.
Galeria de imagens


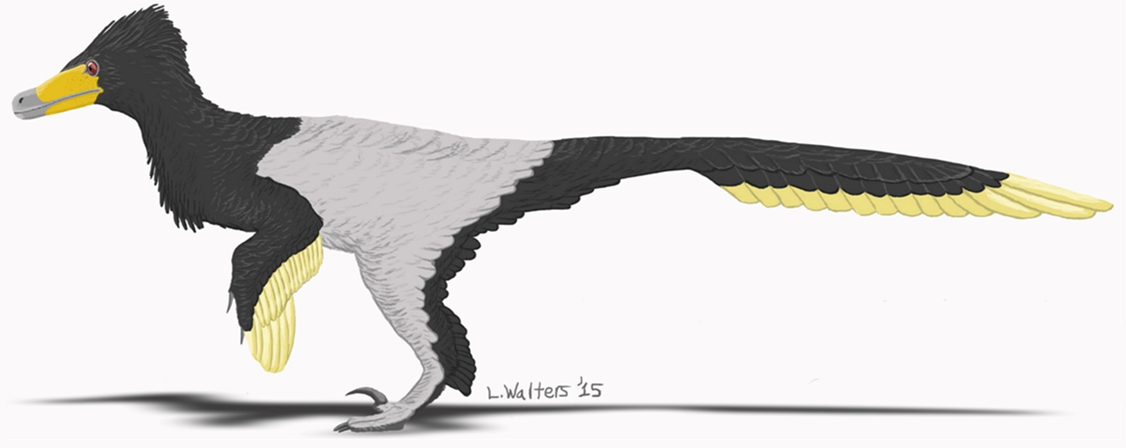









Restauração científica de Velociraptor mongoliensis por Fred Wierum (2017), mostrando cobertura de penas completa, postura horizontal correta e garra falciforme elevada no segundo dedo do pé.
Fred Wierum — CC BY-SA 4.0
Ecologia e comportamento
Habitat
O Velociraptor mongoliensis habitava os ambientes áridos e semiáridos da Formação Djadochta, no atual deserto de Gobi, Mongólia, há 75-71 milhões de anos. O ambiente era dominado por dunas eólicas com ventos constantes, correntes fluviais intermitentes e vegetação esparsa de arbustos e plantas resistentes à seca. As temperaturas eram mais elevadas que as atuais da região. A fauna associada incluía Protoceratops andrewsi, Oviraptor philoceratops, Pinacosaurus grangeri, Zalambdalestes (mamífero) e vários lagartos. O ambiente era muito diferente das selvas exuberantes mostradas em Jurassic Park.
Alimentação
Velociraptor era um predador ativo de pequenos vertebrados: lagartos, pequenos mamíferos, ovos e possivelmente Protoceratops jovens. O espécime dos Dinossauros Lutadores demonstra que atacava presas menores ou iguais ao seu tamanho, usando a garra falciforme para prender e o peso corporal para derrubar a presa — não para evisceração, como indicam análises biomecânicas recentes. Os dentes recurvados e serrilhados eram adaptados para cortar carne. Evidências de dentes de Velociraptor associados a ossos marcados de Protoceratops (Hone et al. 2010) sugerem que também consumia carcaças de animais maiores.
Comportamento e sentidos
As evidências diretas de comportamento incluem o espécime dos Dinossauros Lutadores (predação) e knobs de cálamos (penas para possível mantling ou exibição). Ao contrário da representação em Jurassic Park, não há evidências de caça em grupo coordenado — este comportamento é extrapolação. A presença de penas sugere possível comportamento de aquecimento de ninhos análogo ao de aves modernas. Os bulbos olfatórios e o labirinto do ouvido interno, analisados por King et al. (2020), indicam olfação e audição aguçadas, consistentes com caçador crepuscular ou noturno no ambiente desértico.
Fisiologia e crescimento
Velociraptor era quase certamente endotérmico (de 'sangue quente'), com metabolismo elevado similar ao de aves modernas. A histologia óssea de dromeossaurídeos mostra tecido ósseo fibrolamelar com crescimento rápido, indicativo de endotermia. A presença de penas, confirmada pelos knobs de cálamos, servia primariamente para termorregulação (isolamento) no ambiente árido com grandes variações térmicas diárias. O tamanho pequeno (~20 kg) permitia metabolismo ativo e possivelmente acelerado. A estrutura cerebral revelada por micro-CT (King et al. 2020) é mais próxima a aves do que a répteis em termos de proporções relativas dos centros sensoriais.
Paleogeografia
Configuração continental

Ron Blakey · CC BY 3.0 · Cretáceo, ~90 Ma
Sítios fóssilíferos

Tsogtbaatar, Weishampel, Evans & Watabe — CC BY 2.5
Durante o Campaniano (~75–71 Ma), Velociraptor mongoliensis habitava a Laramídia, a metade ocidental do que hoje é a América do Norte, separada pelo Mar Interior do Oeste (Western Interior Seaway), um mar raso que dividia o continente ao meio. Os continentes estavam em posições muito diferentes das atuais: a Índia viajava em direção à Ásia, a Antártida ainda estava conectada à Austrália, e a América do Sul era uma ilha separada.
Inventário de Ossos
Baseado em múltiplos espécimes. O holótipo AMNH 6515 consiste apenas em crânio e falanges manuais. O espécime MPC-D 100/985, descrito por Norell & Makovicky (1999), é o mais completo e inclui elementos pós-cranianos detalhados como fúrcula, coracóides e pélvis. O famoso espécime dos 'Dinossauros Lutadores' (MPC-D 100/25) preserva crânio e garras in situ.
Estruturas encontradas
Estruturas inferidas
Literatura Científica
15 artigos em ordem cronológica — do artigo de descrição original até pesquisas recentes.
Three new Theropoda, Protoceratops zone, central Mongolia
Osborn, H.F. · American Museum Novitates
Artigo fundador que estabelece o gênero e a espécie Velociraptor mongoliensis. Henry Fairfield Osborn descreve o holótipo AMNH 6515 — um crânio lateralmente comprimido e uma falange ungueal manual — coletado por Peter Kaisen na expedição do American Museum of Natural History às Penhascos Flamejantes (Bayn Dzak), Mongólia, em agosto de 1923. Osborn diagnostica o novo táxon pelos dentes recurvados em série densa, pelo crânio baixo e alongado e pelo tamanho corporal relativamente pequeno comparado a outros terópodes do Cretáceo. O nome mongoliensis refere-se ao país de origem. Publicado como American Museum Novitates 144, é um dos primeiros papers sobre dinossauros mongoleses e lança as bases para um século de pesquisa sobre dromeossaurídeos.
Important features of the dromaeosaur skeleton: information from a new specimen
Norell, M.A. & Makovicky, P.J. · American Museum Novitates
Norell e Makovicky descrevem um novo espécime parcial e tridimensionalmente preservado de dromeossaurídeo (IGM 100/985) coletado em Tugrugeen Shireh, Mongólia, durante as expedições conjuntas AMNH-Academia de Ciências da Mongólia de 1993. O espécime, referido a Velociraptor mongoliensis, documenta pela primeira vez vários elementos pós-cranianos do gênero com qualidade excepcional: fúrcula, placas esternais páreadas com sulcos articulares para os coracóides e escápula em posição subhorizontal em relação à coluna dorsal. Esses caracteres são notavelmente similares aos encontrados em Archaeopteryx e aves basais. Publicado como American Museum Novitates 3215.

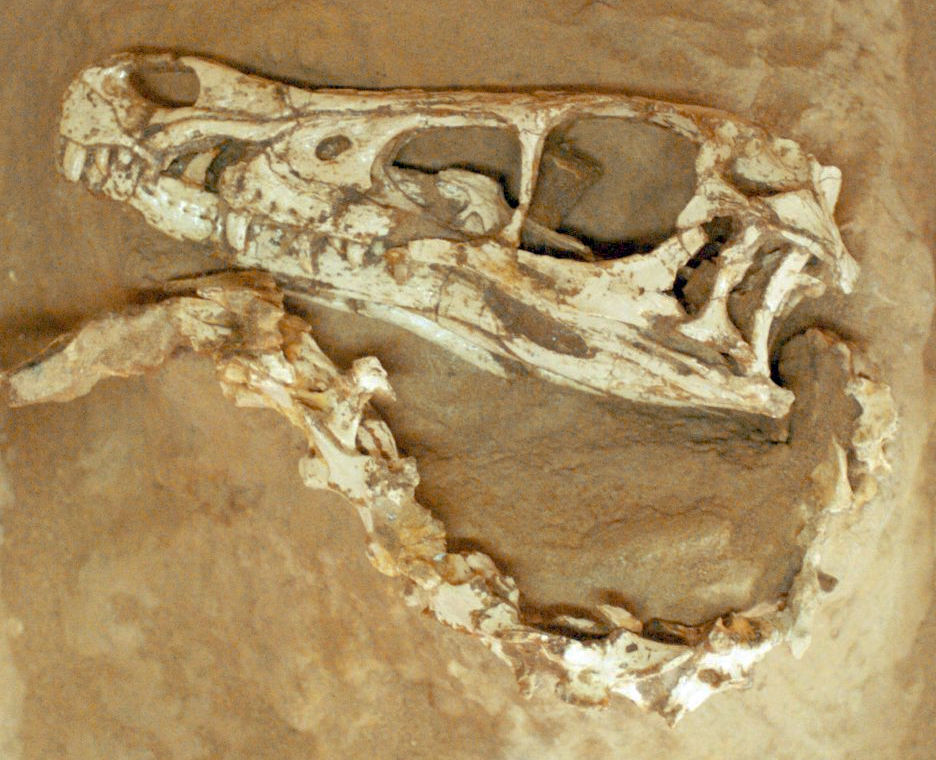
The skull of Velociraptor (Theropoda) from the Late Cretáceous of Mongolia
Barsbold, R. & Osmólska, H. · Acta Palaeontologica Polonica
Barsbold e Osmólska fornecem a análise craniana mais detalhada do Velociraptor mongoliensis até então, examinando material de múltiplos espécimes incluindo o holótipo AMNH 6515 e o espécime dos Dinossauros Lutadores (MPC-D 100/25). O trabalho documenta sistemáticamente as diferenças cranianas que distinguem Velociraptor de Deinonychus: a arcada supratemporal lateralmente convexa, o nasal deprimido, o processo maxilar mais longo e a borda ventral convexa do dentário. Publicado em Acta Palaeontologica Polonica vol. 44(2): 189-219, é a referência anatômica primária para morfologia craniana do táxon.
Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis
Norell, M.A. & Makovicky, P.J. · American Museum Novitates
Segunda parte da série monográfica de Norell e Makovicky sobre anatomia pós-craniana de Velociraptor mongoliensis, publicada como American Museum Novitates 3282, com 45 páginas. O trabalho descreve múltiplos espécimes recém-coletados e concentra-se nos elementos esqueleticos menos conhecidos: cinto peitoral, membro anterior e cintura pélvica. A descoberta central é que Velociraptor apresenta vários caracteres pós-cranianos quasi-avianos: prateleira anti-ilíaca reduzida, fúrcula, escápula em posição subhorizontal em relação à coluna dorsal e placas esternais que articulam com os coracóides. Esses caracteres aproximam morfologicamente o Velociraptor de Archaeopteryx, reforçando a posição dos dromeossaurídeos como parentes próximos das aves.
New evidence on deinonychosaurian dinosaurs from the Late Cretáceous of Patagonia
Novas, F.E. & Pol, D. · Nature
Novas e Pol descrevem novo material de deinoniquossauriano do Cretáceo tardio da Patagônia e realizam análise filogenética abrangente de Deinonychosauria. O resultado confirma um clado Velociraptorinae distinto incluindo Velociraptor mongoliensis, Deinonychus antirrhopus e material gondwanano referido. Publicado em Nature 433: 858-861, o trabalho é fundamental para entender a distribuição biogeográfica dos Velociraptorinae e demonstra que a linhagem alcançou tanto o Hemisfério Norte quanto o Sul no Cretáceo tardio. A análise filogenética de Novas & Pol (2005) serviu como referência para estudos sistemáticos subsequentes sobre dromeossaurídeos.
Feather quill knobs in the dinosaur Velociraptor
Turner, A.H., Makovicky, P.J. & Norell, M.A. · Science
Artigo de impacto publicado em Science que fornece a primeira evidência osteológica direta de penas em Velociraptor mongoliensis. Turner, Makovicky e Norell documentam seis knobs de cálamos no antebraço posterior (ulna) do espécime IGM 100/981 da Formação Djadochta. O espaçamento dos knobs preservados sugere que aproximadamente 14 penas secundárias ('penas de voo' do antebraço) estavam presentes — número comparável ao de Archaeopteryx. Este achado coloca o Velociraptor entre os dromeossaurídeos confirmadamente emplumados, reforçando a transição evolutiva dinossauros-aves. A descoberta também levanta questão funcional: se Velociraptor tinha penas de voo mas não voava, qual era a função? Hipóteses incluem termorregulação, exibição, controle de manobra ao perseguir presas e proteção de ninhos.
A basal dromaeosaurid and size evolution preceding avian flight
Turner, A.H., Pol, D., Clarke, J.A., Erickson, G.M. & Norell, M.A. · Science
Turner et al. descrevem Mahakala omnogovae, um novo dromeossaurídeo basal do Cretáceo tardio da Mongólia, e realizam análise filogenética abrangente de Paraves. Mahakala é o mais basal dos dromeossaurídeos conhecidos, e sua posição filogenética, combinada com dados de tamanho em todo o grupo, indica que a miniaturização extrema era ancestral em Paraves — ou seja, o ancestral comum de aves, troodontídeos e dromeossaurídeos era pequeno. Velociraptor mongoliensis é recuperado dentro de Velociraptorinae. Publicado em Science 317: 1378-1381, o trabalho é fundamental para entender a evolução do voo aviário e a posição exata de Velociraptor na árvore filogenética dos dinossauros plumados.


Biomechanics of dromaeosaurid dinosaur claws: application of X-ray microtomography, nanoindentation, and finite element analysis
Manning, P.L. et al. · The Anatômical Record
Manning et al. aplicam uma combinação inovadora de técnicas ao estudo das garras falciformes dos dromeossaurídeos: microtomografia de raios-X (micro-CT), nanoindentação e análise de elementos finitos (FEA). O resultado contradiz a hipótese de que a garra falciforme era usada para evisceração de presas grandes. O modelo FEA confirma que as garras eram bem adaptadas para escalada: resistentes a forças no plano longitudinal, mas limitadas em forças tangenciais. A ponta da garra funcionava como elemento de punção e fixação, enquanto a porção proximal expandida transferia estresse de carga através de trabéculas e osso cortical. Publicado em The Anatômical Record 292(9): 1397-1405, este é um dos primeiros estudos a aplicar FEA a material paleontológico de dromeossaurídeos.


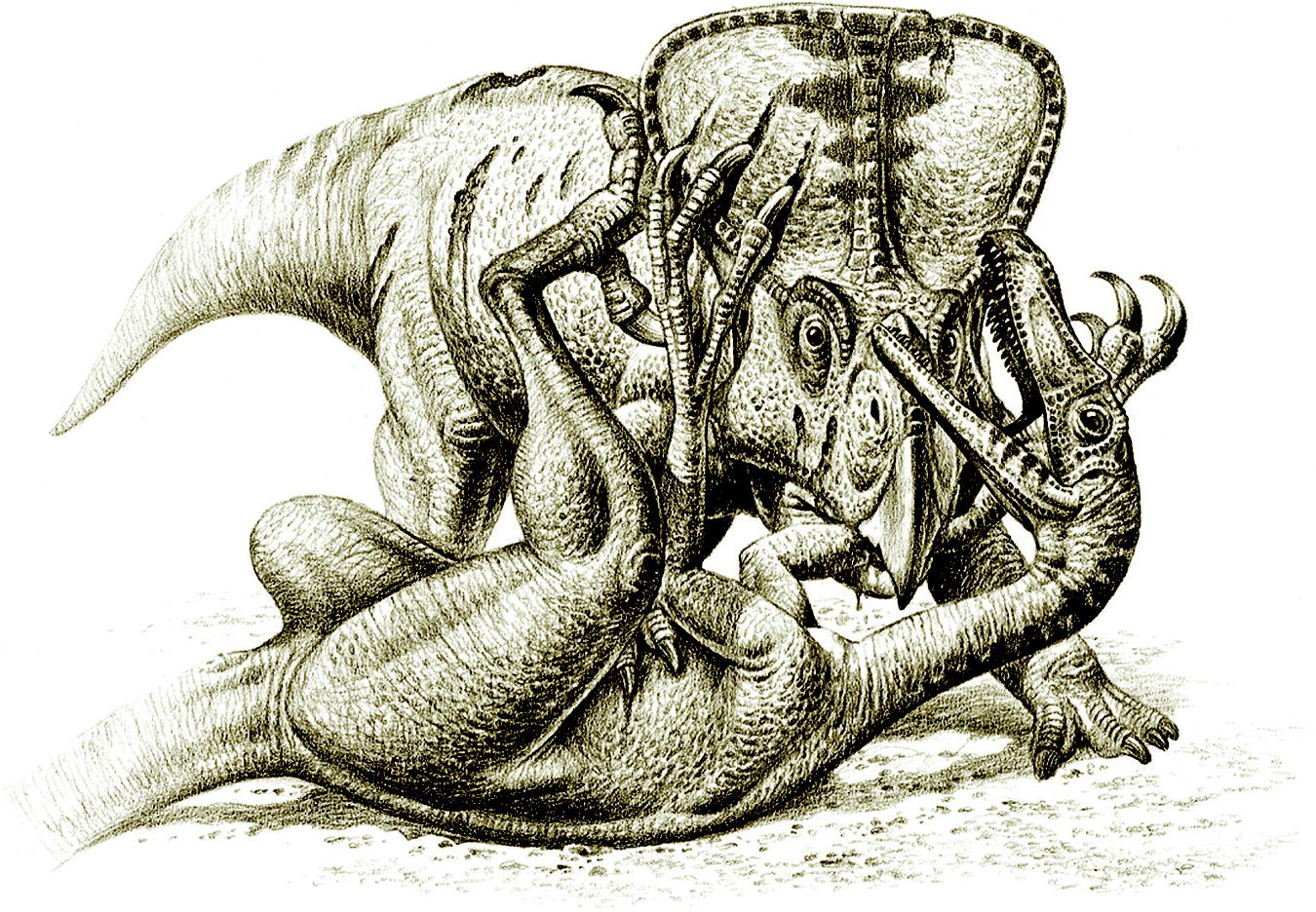
Evidence of predatory behavior by carnivorous dinosaurs
Carpenter, K. · Gaia
Carpenter analisa o espécime dos Dinossauros Lutadores (MPC-D 100/25) como evidência conclusiva de comportamento predatório ativo em terópodes. O espécime, descoberto por uma expedição polonesa-mongol em 1971 na Formação Djadochta, preserva um Velociraptor mongoliensis com a garra falciforme próxima à região jugular de Protoceratops andrewsi, enquanto o ceratopsiano mordia e esmagava o antebraço do predador. Carpenter interpreta a cena como um ataque de predação interrompido por um deslizamento de areia que enterrou ambos os animais vivos. O trabalho foi publicado em Gaia 15: 135-144 e é a referência primária para interpretações comportamentais baseadas no espécime dos Dinossauros Lutadores.
Late Mesozoic stratigraphy and vertebrates of the Gobi Basin
Jerzykiewicz, T. & Russell, D.A. · Cretáceous Research
Jerzykiewicz e Russell apresentam a compilação mais abrangente da estratigrafia do Cretáceo mongol, publicada em Cretáceous Research 12: 345-377. O trabalho documenta como a região do Gobi foi gradualmente transformada em ambiente desértico semiárido durante o Campaniano, representado pelas formações Djadochta e Baruungoyot. A evidência sedimentológica revela um mosaico complexo de fácies: deposição de leques aluviais, fluviais de curta duração e áreas lacustres menores. Os autores correlacionam as faunas de vertebrados de diferentes formações e fornecem o contexto paleoambiental essencial para entender a ecologia de Velociraptor: um ambiente árido de dunas eólicas com recursos de água escassos e uma fauna associada incluindo Protoceratops, Oviraptor e pequenos mamíferos.

New evidence for a trophic relationship between the dinosaurs Velociraptor and Protoceratops
Hone, D., Choiniere, J., Sullivan, C., Xu, X., Pittman, M. & Tan, Q. · Palaeogeography, Palaeoclimatology, Palaeoecology
Hone et al. reportam nova evidência de relação trófica entre Velociraptor e Protoceratops, complementando o espécime dos Dinossauros Lutadores. Na Formação Bayan Mandahu (Mongólia Interior, China), foram encontrados um dente de Velociraptor e ossos de Protoceratops com marcas de dentes compatíveis, em associação. O trabalho é publicado em Palaeogeography, Palaeoclimatology, Palaeoecology 291: 488-492 e é importante por demonstrar que a relação predador-presa entre Velociraptor e Protoceratops não era evento único, mas padrão ecológico documentável em múltiplas localidades e formações. A análise das marcas de dentes nos ossos de Protoceratops indica processamento de carcaça, sugerindo que Velociraptor tanto caçava quanto consumia presas já mortas.
A review of dromaeosaurid systematics and paravian phylogeny
Turner, A.H., Makovicky, P.J. & Norell, M.A. · Bulletin of the American Museum of Natural History
A monografia mais abrangente sobre sistemática de dromeossaurídeos, publicada no Bulletin of the American Museum of Natural History 371: 1-206. Turner, Makovicky e Norell revisam todos os 31 táxons de Dromaeosauridae nomeados até então, validando 26 deles com base em diagnoses fundamentadas em apomorfias. A análise filogenética de Paraves é a maior já realizada para o grupo. Velociraptor mongoliensis é confirmado como espécie-tipo de Velociraptorinae. O trabalho inclui descrições osteológicas detalhadas, matrizes de dados com centenas de caracteres e discussão das relações biogeográficas. Esta monografia é a referência taxonômica fundamental para Dromaeosauridae e o ponto de partida obrigatório para qualquer estudo filogenético posterior do grupo.


A new species of Velociraptor (Dinosauria: Dromaeosauridae) from the Upper Cretáceous of northern China
Godefroit, P., Currie, P.J., Li, H., Shang, C.Y. & Dong, Z. · Journal of Vertebrate Paleontology
Godefroit et al. descrevem Velociraptor osmolskae n. sp. com base em maxilas páreadas e um lacrimal esquerdo (holótipo IMM 99NM-BYM-3/3) da Formação Bayan Mandahu, Mongólia Interior, China — uma segunda formação correlata à Djadochta. Publicado em Journal of Vertebrate Paleontology 28(2): 432-438, este trabalho é importante por expandir o gênero Velociraptor para a China e por documentar variação morfológica intraespecífica ou interespecífica dentro do gênero. O holótipo foi coletado durante uma expedição sino-belga em 1999. A espécie é nomeada em homenagem à paleontóloga polonesa Halszka Osmólska, falecida em março de 2008.


The endocranium and trophic ecology of Velociraptor mongoliensis
King, J.L., Sipla, J.S., Georgi, J.A., Balanoff, A.M. & Neenan, J.M. · Journal of Anatomy
King et al. aplicam microtomografia computadorizada de raios-X (micro-CT) para reconstruir digitalmente a anatomia endocraniana de Velociraptor mongoliensis, publicando em Journal of Anatomy 237(5): 861-869. A reconstrução inclui o endocast cerebral, nervos cranianos, estruturas vasculares e o labirinto endósseo do ouvido interno. O resultado revela que Velociraptor possuía bulbos olfatórios relativamente grandes (indicando olfação aguda), cerebelo desenvolvido (indicando bom equilíbrio e coordenação) e canais semicirculares do labirinto com geometria compatível com movimentos cefálicos rápidos. Esses dados neuroanatômicos são consistentes com um predador ativo altamente sensorial, capaz de detectar presas a distância e realizar ataques precisos.


Velociraptor: the state of the art 100 years after the discovery of this iconic dinosaur
Bindellini, G., Moscarella, A., Makovicky, P., Manucci, F. & Romano, M. · Italian Journal of Geosciences
Artigo de revisão comemorativo do centenário da descrição de Velociraptor mongoliensis, publicado em Italian Journal of Geosciences 144(3): 460-486. Bindellini et al. apresentam uma nova reconstrução esquelética do espécime MUST SN1140/BM digitalizado por fotogrametria, e sintetizam o estado da arte de 100 anos de pesquisa. O trabalho abrange anatomia, filogenia, comportamento, paleoecologia e a evolução das interpretações científicas sobre o animal. É a síntese mais atualizada sobre Velociraptor disponível, incorporando dados de micro-CT, análise filogenética moderna e paleoarte baseada em evidências. Publicado com Peter Makovicky como coautor, um dos maiores especialistas em dromeossaurídeos, este artigo é referência atual obrigatória.


Espécimes famosos em museus
AMNH 6515 (Holótipo)
American Museum of Natural History, New York, EUA
Holótipo de Velociraptor mongoliensis, consistindo em crânio lateralmente comprimido e uma falange ungueal manual, coletado nas Penhascos Flamejantes (Bayn Dzak) em agosto de 1923. Base da descrição original de Osborn (1924).
MPC-D 100/25 — 'Dinossauros Lutadores'
Museu de Paleontologia da Mongólia, Ulaanbaatar
Espécime mais famoso do mundo: Velociraptor mongoliensis e Protoceratops andrewsi preservados em combate. Descoberto em Tugrik Shire em 1971, é o registro mais direto de comportamento predatório em dinossauros já encontrado e é patrimônio natural da Mongólia.
MPC-D 100/985
Museu de Paleontologia da Mongólia, Ulaanbaatar
Espécime mais completo de Velociraptor mongoliensis com pós-craniano preservado, coletado em Tugrugeen Shireh em 1993. Base do trabalho de Norell & Makovicky (1997, 1999) que documentou fúrcula, placas esternais e outros elementos quasi-avianos.
No cinema e na cultura popular
Poucos dinossauros deixaram marca tão profunda na cultura pop quanto o Velociraptor, graças sobretudo a Jurassic Park (1993). O filme de Steven Spielberg transformou um animal do tamanho de um peru em um monstro de dois metros capaz de abrir portas e coordenar emboscadas, usando a morfologia de Deinonychus em vez do animal real. O resultado foi um ícone cultural permanente. As sequências Lost World (1997), Jurassic Park III (2001) e a franquia Jurassic World (2015-2022) mantiveram e amplificaram essa representação, chegando aos Velociraptores com nomes próprios e treinamento militar de Jurassic World. Do lado oposto, a série Prehistoric Planet (Apple TV+, 2022) com narração de David Attenborough apresentou pela primeira vez a um grande público um Velociraptor emplumado, de tamanho correto e comportamento baseado em evidências, marcando uma virada na representação midiática. A discrepância entre o Velociraptor fílmico e o científico tornou-se ela mesma um tema cultural: paleontólogos como Jack Horner e Mark Norell comentam regularmente sobre os erros de Jurassic Park, e a frase 'o Velociraptor real era do tamanho de um peru' entrou no vocabulário popular da divulgação científica.

Classificação
Descoberta
Curiosidade
O Velociraptor real era aproximadamente do tamanho de um peru — não do tamanho de um humano como mostrado em Jurassic Park. O filme usou a morfologia de Deinonychus, um dromeossaurídeo duas vezes maior que o Velociraptor. Quando Jurassic Park foi lançado em 1993, o Utahraptor — um dromeossaurídeo realmente do tamanho retratado no filme — foi descoberto e descrito, levando o designer Stan Winston a brincar: 'Depois que criamos o nosso, eles o descobriram'.