Rajasauro
Rajasaurus narmadensis
"Lagarto real do Narmada"
Sobre esta espécie
O Rajasaurus narmadensis foi o maior predador terrestre do subcontinente indiano no Cretáceo Superior. Viveu entre 69 e 66 milhões de anos atrás no que hoje é o estado de Gujarat, no noroeste da Índia, em um ambiente árido-semiárido varrido por rios sazonais e marcado pelos episódios vulcânicos das Armadilhas do Decã. Único abelissaurídeo indiano com restos pós-cranianos bem preservados, destacou-se por uma crista sagital mediana sobre o crânio, característica única entre os abelissaurídeos indianos. Sua classificação dentro de Majungasaurinae confirma a conexão biogeográfica entre a Índia e Madagascar no Cretáceo tardio.
Formação geológica e ambiente
A Formação Lameta é uma unidade sedimentar do Maastrichtiano (Cretáceo Superior, ~69-66 Ma) aflorante em Gujarat, Madhya Pradesh, Maharashtra e estados adjacentes da Índia central e ocidental. Consiste em alternâncias de argilitos, siltitos e arenitos depositados em condições fluviais e lacustres em clima semiárido a tropical úmido-seco. A formação é internacionalmente reconhecida por sua extraordinária riqueza de ninhos e ovos de sauropodas titanossaurídeos (nove ooespécies descritas), além de restos de abelissaurídeos como o Rajasaurus, Rahiolisaurus e Indosuchus. Deposited imediatamente abaixo dos basaltos das Armadilhas do Decã, a formação marca os últimos ambientes terrestres antes da grande extinção do Cretáceo-Paleógeno na região.
Galeria de imagens
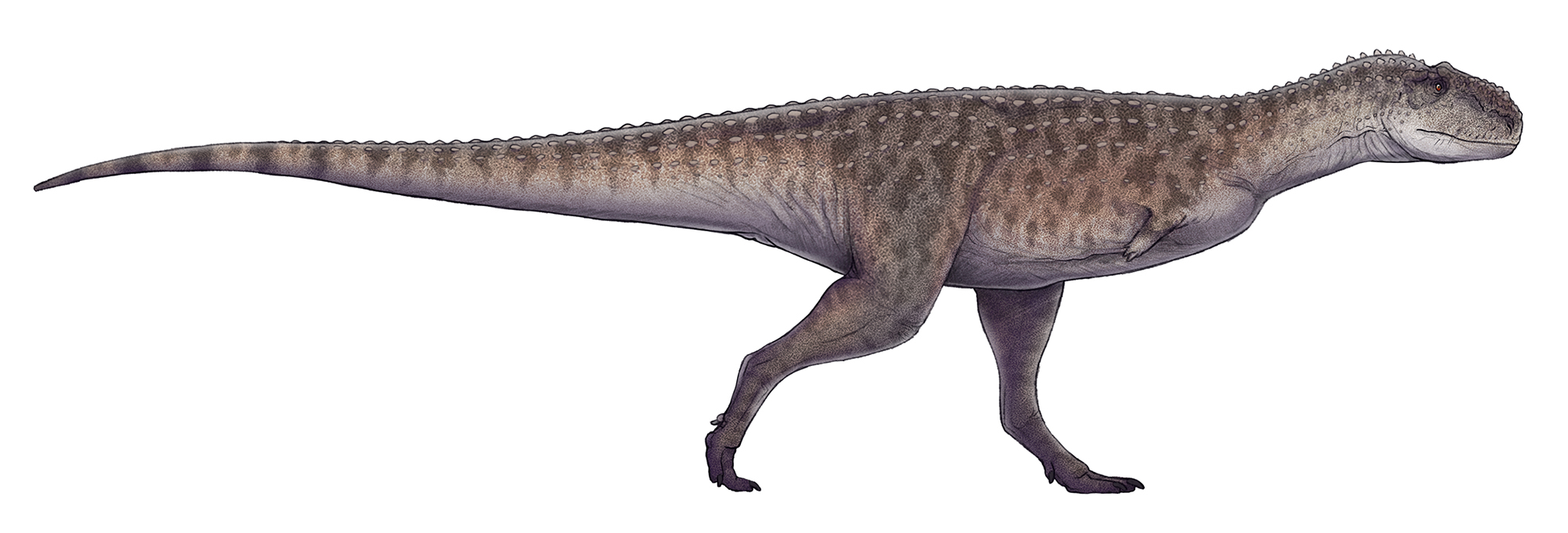













Reconstituição de vida do Rajasaurus narmadensis baseada no holótipo GSI 21141/1-33, por Paleocolour (2017). Vista lateral mostrando as proporções corporais do único abelissaurídeo indiano bem descrito.
Paleocolour, CC BY-SA 3.0
Ecologia e comportamento
Habitat
O Rajasaurus habitou a bacia do Rio Narmada no que hoje é o Gujarat, no noroeste da Índia, durante o Maastrichtiano tardio (69-66 Ma). O ambiente era uma planície aluvial árida a semiárida, com rios sazonais e lagos efêmeros, intercalados por episódios de vulcanismo intenso das emergentes Armadilhas do Decã. A vegetação era dominada por plantas vasculares primitivas, incluindo gramíneas em evolução, samambaias e coníferas. Compartilhava o ecossistema com titanossauros como o Isisaurus colberti, o sauropoda indiano mais bem conhecido.
Alimentação
Predador de topo do ecossistema da Formação Lameta, o Rajasaurus provavelmente caçava titanossauros adultos e subadultos, filhotes recém-eclodidos de ninhos de sauropodas, e potencialmente outros répteis de médio porte. O crânio curto e robusto dos abelissaurídeos sugere mordida voltada para a força no eixo vertical mais do que para corte longitudinal. A musculatura cervical poderosa, inferida da anatomia das vértebras, permitia agarrar e controlar presas grandes. A cena de caça a filhotes de Isisaurus no Deccan, reconstruída em Prehistoric Planet, é biologicamente plausível.
Comportamento e sentidos
Evidências diretas de comportamento são escassas dado o registro fóssil limitado do Rajasaurus. A crista mediana sobre o crânio pode ter funcionado para reconhecimento intraespecífico ou como estrutura de display em interações entre indivíduos — hipótese proposta por Delcourt (2018) para abelissaurídeos em geral. A presença de pelo menos dois outros abelissaurídeos (Rahiolisaurus e Indosuchus) na mesma formação geológica sugere possível estratificação de nicho ou diferenciação de presas entre os predadores coexistentes.
Fisiologia e crescimento
Como abelissaurídeo derivado, o Rajasaurus provavelmente apresentava metabolismo elevado semelhante ao de outros grandes terópodes do Cretáceo, com crescimento relativamente rápido inferido por comparação com parentes bem estudados como o Majungasaurus. Os membros anteriores eram extremamente reduzidos, como em todos os abelissaurídeos, e provavelmente não participavam ativamente da predação. O crânio relativamente curto e alto, com musculatura cervical poderosa, sugere que o pescoço e a cabeça eram as ferramentas primárias de ataque e manipulação de presas.
Paleogeografia
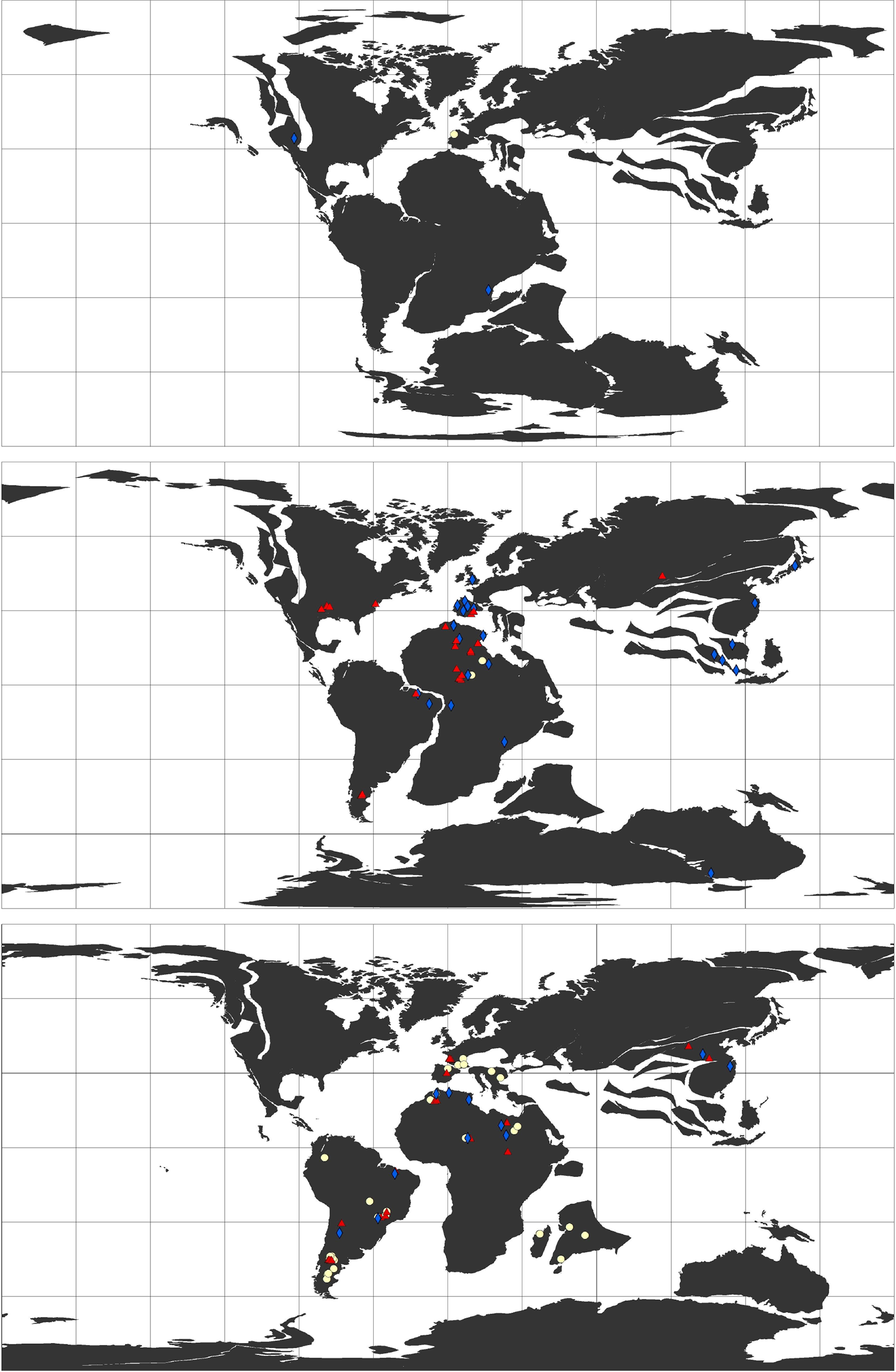
Configuração continental

Ron Blakey · CC BY 3.0 · Cretáceo, ~90 Ma
Durante o Maastrichtiano (~69–66 Ma), Rajasaurus narmadensis habitava a Laramídia, a metade ocidental do que hoje é a América do Norte, separada pelo Mar Interior do Oeste (Western Interior Seaway), um mar raso que dividia o continente ao meio. Os continentes estavam em posições muito diferentes das atuais: a Índia viajava em direção à Ásia, a Antártida ainda estava conectada à Austrália, e a América do Sul era uma ilha separada.
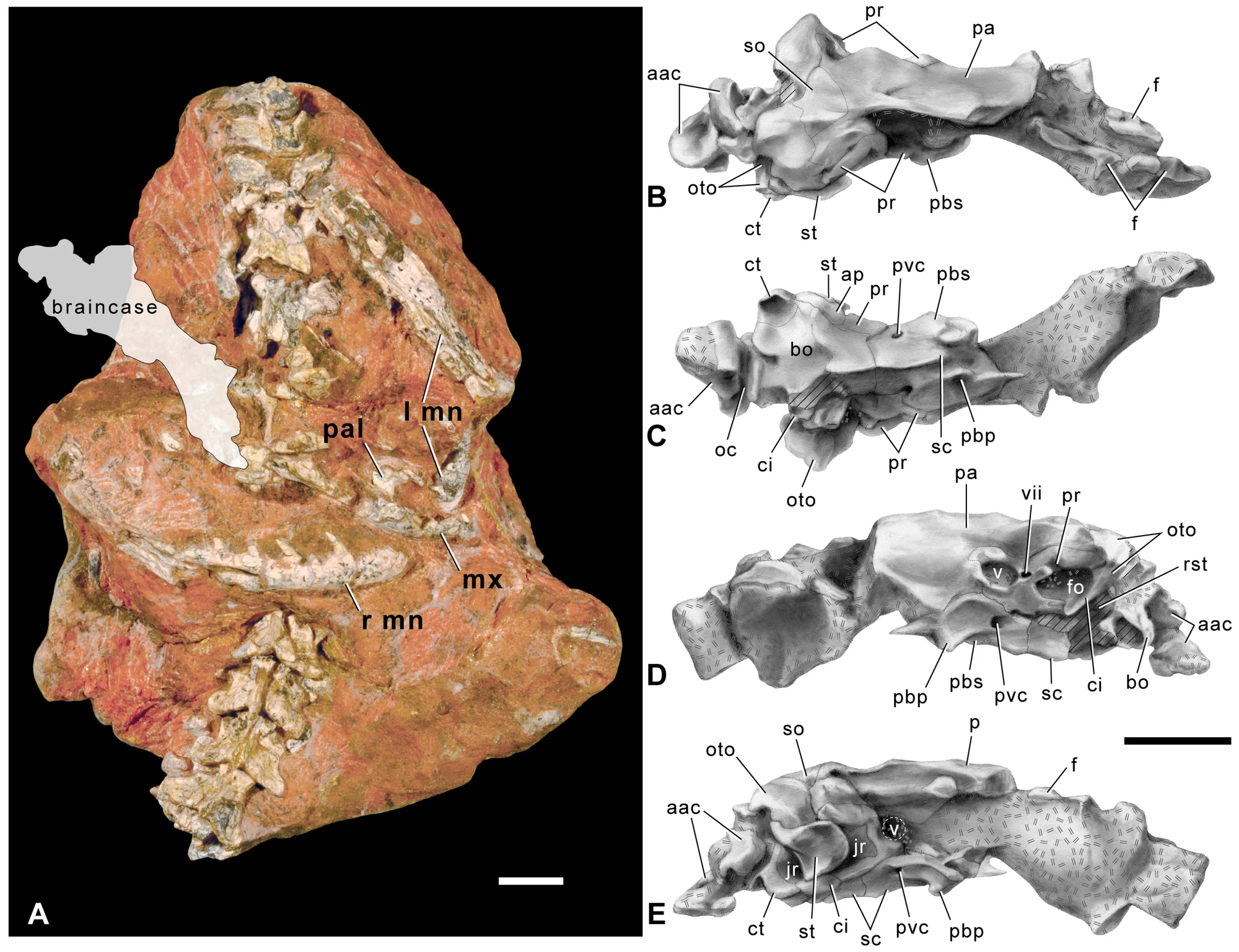
Inventário de Ossos
O holótipo GSI 21141/1-33 preserva maxilas, pré-maxilas, braincase, quadrado, uma vértebra cervical, 11 vértebras dorsais parciais, seis sacrais, três caudais, o ílio, fíbula e metatarsos. É o único abelissaurídeo indiano com restos pós-cranianos significativos, o que o torna extremamente valioso apesar da incompletude.
Estruturas encontradas
Estruturas inferidas
Literatura Científica
15 artigos em ordem cronológica — do artigo de descrição original até pesquisas recentes.
A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India
Wilson, J.A., Sereno, P.C., Srivastava, S., Bhatt, D.K., Khosla, A. & Sahni, A. · Contributions from the Museum of Paleontology, University of Michigan
Artigo fundador que descreve formalmente o Rajasaurus narmadensis com base no holótipo GSI 21141/1-33, escavado da Formação Lameta em Gujarat. Wilson, Sereno e colaboradores identificam o animal como o primeiro terópode indiano com restos pós-cranianos bem preservados, o que permitiu uma caracterização anatômica sem precedente para terópodes indianos. O crânio preserva a crista sagital mediana, formada pelos ossos frontal e nasal, característica compartilhada com o Majungasaurus de Madagascar. O trabalho estabelece a posição filogenética do Rajasaurus nos Carnotaurinae, sugerindo conexão biogeográfica entre a Índia e Madagascar antes da separação final das massas continentais. A diagnose formal e a comparação com outros abelissaurídeos gondwânicos tornaram este paper a referência primária para qualquer estudo sobre o gênero.
The phylogeny of Ceratosauria (Dinosauria: Theropoda)
Carrano, M.T. & Sampson, S.D. · Journal of Systematic Palaeontology
Análise filogenética abrangente de toda a Ceratosauria, incluindo todos os táxons principais conhecidos até 2008. Carrano e Sampson revisam os caracteres diagnósticos do grupo e apresentam uma topologia revisada que posiciona o Rajasaurus firmemente dentro dos Abelisauridae, como parente próximo do Majungasaurus de Madagascar. O estudo mapeia a distribuição geográfica e temporal dos ceratossauros, confirmando a presença desses predadores em todos os continentes do Gondwana. A análise inclui caracteres cranianos e pós-cranianos, o que permite integrar o Rajasaurus, com seus elementos pós-cranianos parcialmente preservados, na matriz de dados de forma mais robusta do que estudos anteriores.
Rahiolisaurus gujaratensis, n. gen. n. sp., a new abelisaurid theropod from the Late Cretaceous of India
Novas, F.E., Chatterjee, S., Rudra, D.K. & Datta, P. · New Aspects of Mesozoic Biodiversity (Lecture Notes in Earth Sciences, vol. 132)
Descrição formal de um novo abelissaurídeo da Formação Lameta, o Rahiolisaurus gujaratensis, escavado próximo à mesma localidade de Rahioli onde o Rajasaurus foi encontrado. Novas e colaboradores descrevem restos de pelo menos sete indivíduos, tornando o Rahiolisaurus um dos abelissaurídeos indianos mais bem documentados. O paper é fundamental para entender a diversidade de abelissaurídeos na Índia do Cretáceo: Rajasaurus e Rahiolisaurus coexistiam no mesmo ecossistema, sugerindo nicho diferenciado ou especialização temporal distinta entre os dois predadores. A análise filogenética do estudo, comparando ambos os gêneros, reforça a conexão entre os abelissaurídeos indianos e seus parentes gondwânicos na América do Sul e Madagascar.

Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers
Delcourt, R. · Scientific Reports
Estudo abrangente sobre a paleobiologia dos ceratossauros que inclui análise anatômica e filogenética específica dos abelissaurídeos do hemisfério sul. Delcourt examina especificamente que tanto o Majungasaurus quanto o Rajasaurus narmadensis possuem uma crista mediana única, formada por ossos diferentes em cada gênero (frontal no Majungasaurus, nasofrontal no Rajasaurus), com implicações para a reconstituição dos tecidos moles do crânio. O paper propõe que abelissaurídeos possuíam tecidos moles especializados na superfície craniana, possivelmente relacionados ao reconhecimento intraespecífico ou display. O estudo descreve dois planos corporais principais nos ceratossauros — Noasauridae e Etrigansauria (Ceratosauridae + Abelisauridae) — e situa o Rajasaurus dentro de um padrão biogeográfico gondwânico de longa duração.

Allometry and body length of abelisauroid theropods: Pycnonemosaurus nevesi is the new king
Grillo, O.N. & Delcourt, R. · Cretaceous Research
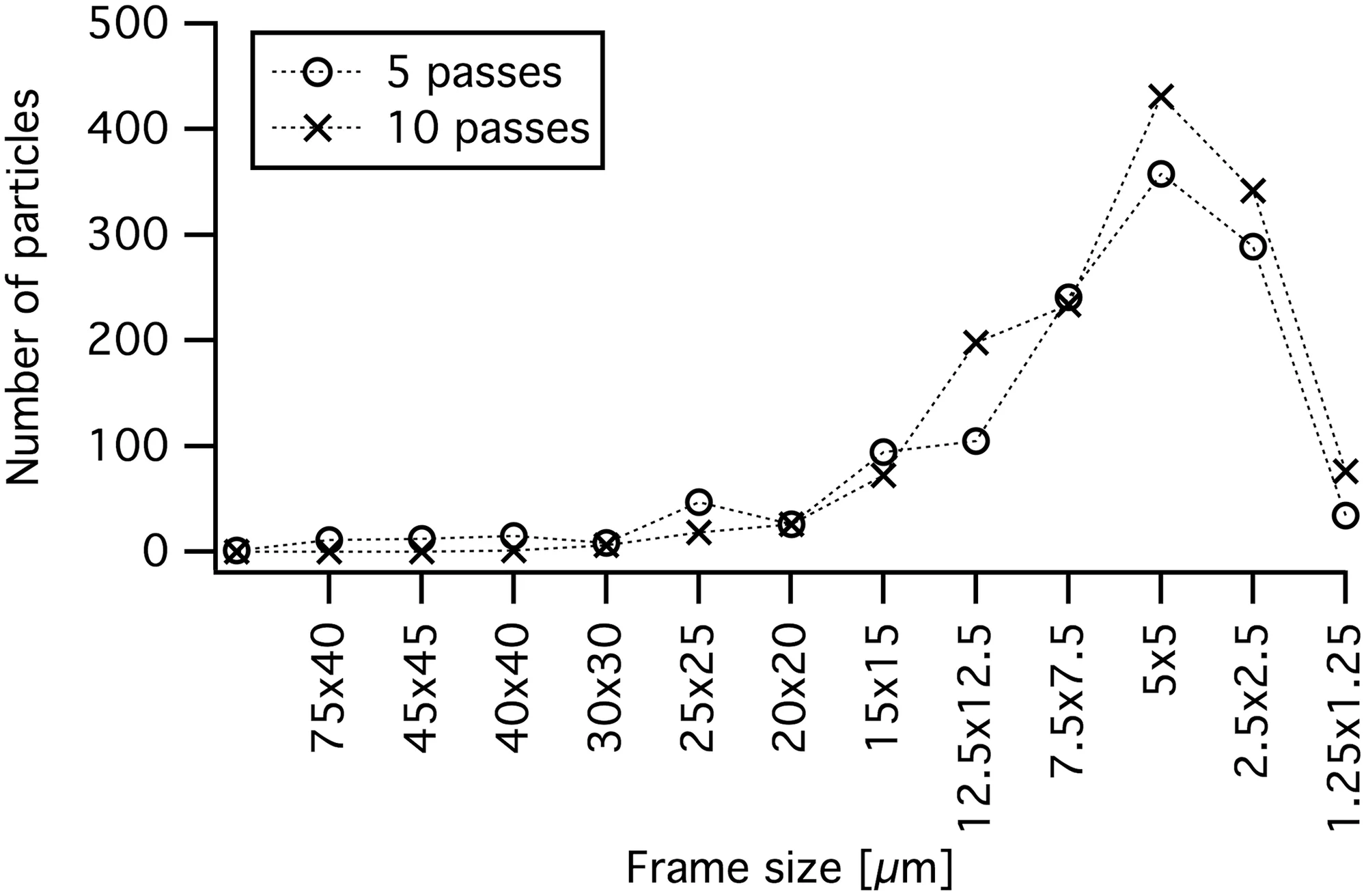
Estudo alométrico sistemático que reavalia as estimativas de comprimento corporal para todos os abelissauroides com base em correlações robustas entre elementos esqueléticos e dimensões corporais totais. Grillo e Delcourt utilizam bivariar equações e 40 medidas do crânio, vértebras e elementos apendiculares para estimar o comprimento corporal do Rajasaurus narmadensis com base nos elementos preservados. O estudo revisa para baixo estimativas anteriores do Rajasaurus (que chegavam a 11 metros em publicações populares), estabelecendo uma estimativa mais conservadora de 6,6 metros baseada na correlação entre o tamanho do fêmur e o comprimento total. Esta revisão alométrica é a base quantitativa mais sólida disponível para o porte corporal do Rajasaurus.
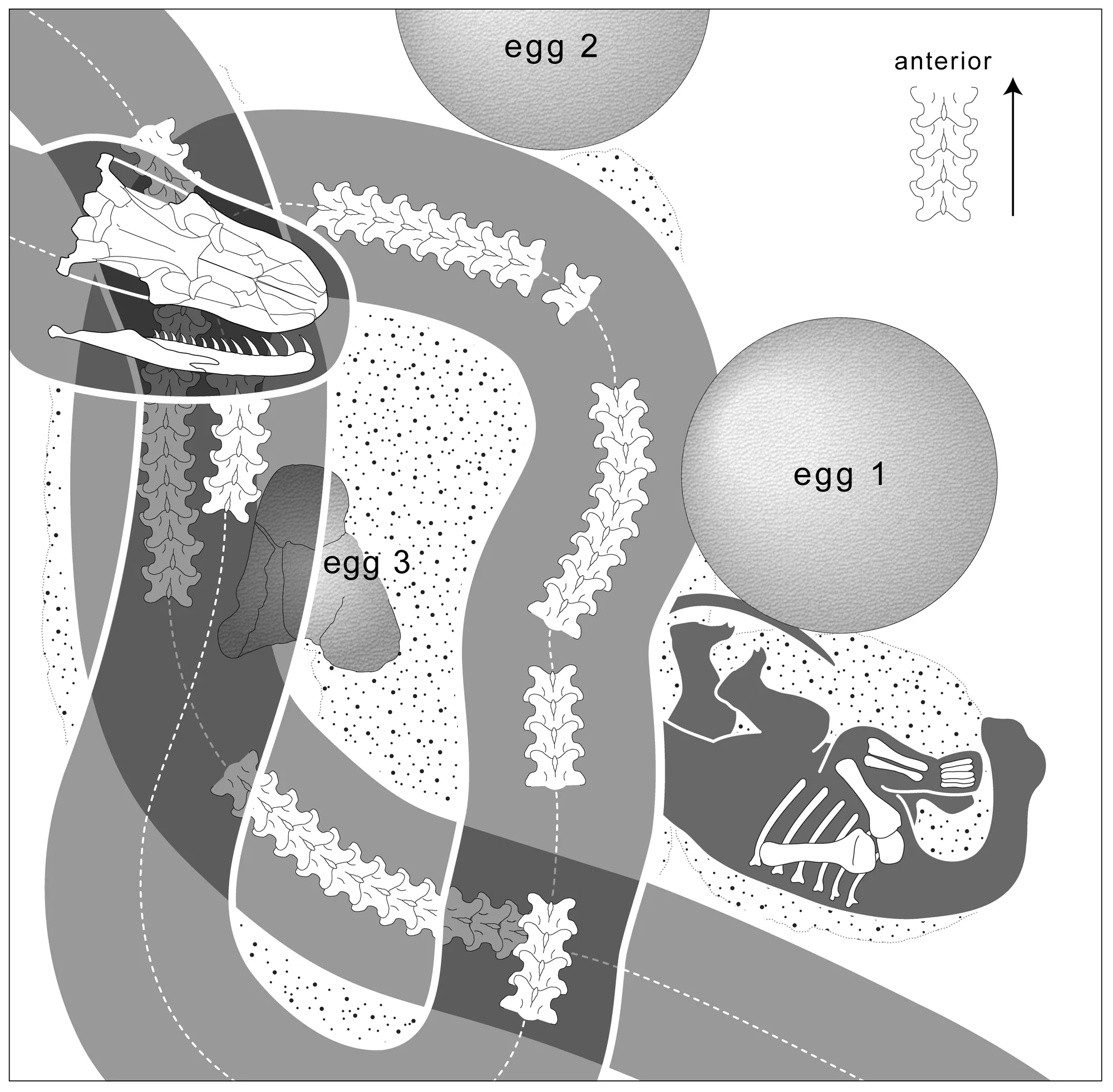
Predation upon hatchling dinosaurs by a new snake from the Late Cretaceous of India
Wilson, J.A., Mohabey, D.M., Peters, S.E. & Head, J.J. · PLOS Biology
Descrição de uma cobra nova de 3,5 metros do Maastrichtiano da Índia (Sanajeh indicus), preservada extraordinariamente dentro de um ninho de sauropoda, enrolada ao redor de um ovo ao lado dos restos de um filhote. O paper documenta predação por um réptil não-dinosauriano sobre filhotes de sauropodas na Formação Lameta — o mesmo ecossistema habitado pelo Rajasaurus. Esta descoberta ilumina a dinâmica ecológica do ambiente cretáceo indiano: os ninhos de titanossauros que Isisaurus colocava às margens dos rios estavam sujeitos a predadores múltiplos, tanto terópodes como o Rajasaurus quanto répteis oportunistas como a cobra Sanajeh. O estudo de Wilson et al. (2010) é fundamental para entender o contexto paleobiológico em que o Rajasaurus caçava.

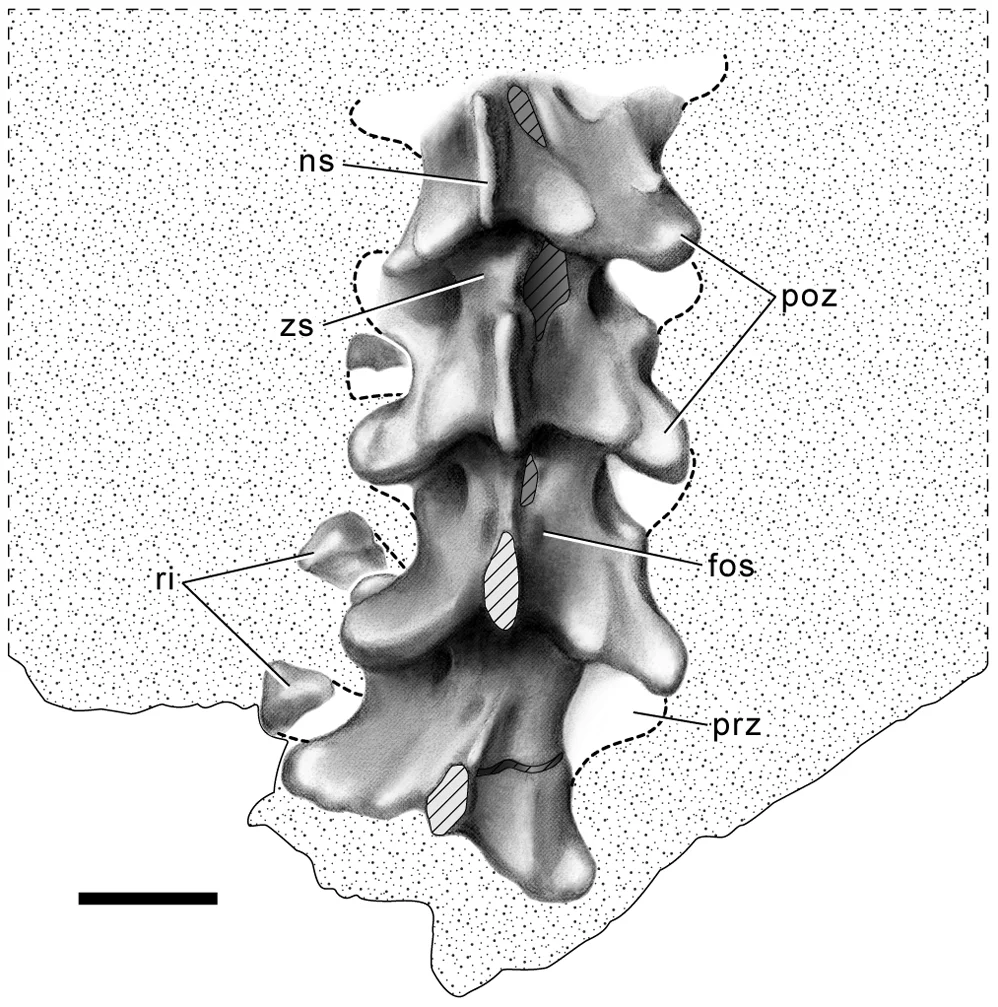
Palaeoenvironments of the dinosaur-bearing Lameta Beds (Maastrichtian), Narmada Valley, Central India
Tandon, S.K., Sood, A., Andrews, J.E. & Dennis, P.F. · Palaeogeography, Palaeoclimatology, Palaeoecology
Análise sedimentológica e geoquímica abrangente dos Leitos Lameta, a unidade estratigráfica que engloba a Formação Lameta onde o Rajasaurus foi encontrado. Tandon e colaboradores identificam quatro unidades mapeáveis na região de Jabalpur: Arenito Verde basal (depósito de riacho entrelaçado), Calcário Inferior (planície palustrina exposta subaerialmente), Leitos Nodulares Mosqueados (depósitos de planície aluvial semiárida pedogenicamente modificados) e Arenito Superior (depósito de inundação laminar). As análises isotópicas de carbono e oxigênio confirmam ambientes de zona de solo terrestre. O quadro reconstituído é de uma planície aluvial árida a semiárida com rios sazonais — o habitat real do Rajasaurus, que caçava neste ambiente varrido por episódios de vulcanismo do Decã.

Microflora from sauropod coprolites and associated sediments of Late Cretaceous (Maastrichtian) Lameta Formation of Nand-Dongargaon basin, Maharashtra
Sonkusare, H., Samant, B. & Mohabey, D.M. · Journal of the Geological Society of India
Estudo palinológico de coprólitos de sauropoda e sedimentos associados da Formação Lameta na bacia de Nand-Dongargaon em Maharashtra, revelando a composição da flora consumida pelos sauropodas herbívoros que viveram no mesmo ecossistema do Rajasaurus. A análise documenta pólen, esporos, restos de algas, fungos e cutículas bem preservadas de Poaceae, além de amebas testáceas. Esta composição sugere que os sauropodas da Formação Lameta consumiam gramináceas primitivas e outras plantas vasculares, dentro de um ambiente semi-árido. Compreender a dieta dos sauropodas é essencial para entender a ecologia do Rajasaurus: eram esses herbívoros as presas potenciais do abelissaurídeo indiano.
Cretaceous theropods from India: a review of specimens described by Huene and Matley (1933)
Novas, F.E., Agnolin, F. & Bandyopadhyay, S. · Revista del Museo Argentino de Ciencias Naturales
Revisão dos espécimes históricos de terópodes cretáceos da Índia descritos originalmente por Huene e Matley em 1933. Novas, Agnolin e Bandyopadhyay reanalisam materiais de Indosaurus matleyi, Indosuchus raptorius, Laevisuchus indicus e outros táxons históricos à luz da anatomia abelissaurídea moderna. O trabalho contextualiza o Rajasaurus narmadensis dentro de uma fauna de abelissaurídeos indianos mais diversa do que anteriormente reconhecida, estabelecendo o cenário taxonômico para entender o papel ecológico desses predadores na Formação Lameta. A reassignação de vários espécimes históricos e a análise de seus caracteres diagnósticos são fundamentais para entender a diversidade de terópodes coexistindo com o Rajasaurus.


Paleoenvironment and paleoecology of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar
Rogers, R.R., Krause, D.W., Curry Rogers, K., Rasoamiaramanana, A.H. & Rahantarisoa, L. · Journal of Vertebrate Paleontology (Memoir 8)
Reconstituição do paleoambiente e paleoecologia do Majungasaurus crenatissimus, o parente mais próximo do Rajasaurus narmadensis, a partir da Formação Maevarano do noroeste de Madagascar. Rogers e colaboradores descrevem condições semiáridas com atividade fluvial sazonal — ambiente notavelmente paralelo ao da Formação Lameta habitada pelo Rajasaurus. Esta comparação é de importância biogeográfica fundamental: Madagascar e Índia compartilhavam não apenas abelissaurídeos aparentados (Majungasaurus e Rajasaurus), mas também condições paleoambientais similares, sugerindo que esses predadores ocupavam ecossistemas ecologicamente equivalentes nos dois landmasses separados. O estudo inclui análise da tafonomia, estratigrafia e fauna associada do Maevarano.

Building a predator: macroevolutionary patterns in the skull of abelisaurid dinosaurs
Pereyra, E.E.S., Ezcurra, M.D., Paschetta, C. & Méndez, A.H. · Proceedings of the Royal Society B: Biological Sciences
Análise macroevolucionária dos padrões de modularidade, integração e disparidade morfológica do crânio de abelissaurídeos, incluindo o Rajasaurus, utilizando morfometria geométrica bidimensional. Pereyra e colaboradores identificam que o neurocrânio foi a principal região responsável pelo aumento proporcional da altura craniana nos abelissaurídeos ao longo da evolução do grupo. Alta disparidade e taxas evolutivas são encontradas nas regiões occipital, esquamosal, quadradojugal, lacrimal e pós-orbital — exatamente as regiões relacionadas à crista craniana do Rajasaurus. O estudo conclui que a especialização alimentar foi provavelmente o principal motor da evolução craniana nos abelissaurídeos, com algumas características cranianas subsequentemente co-optadas para display sociossexual, como a crista mediana do Rajasaurus.


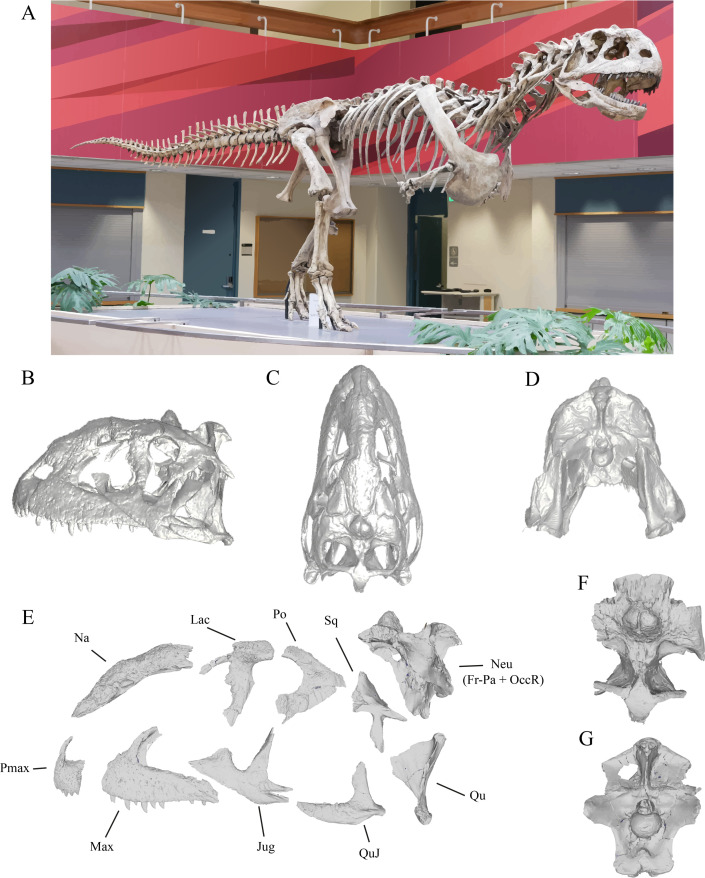
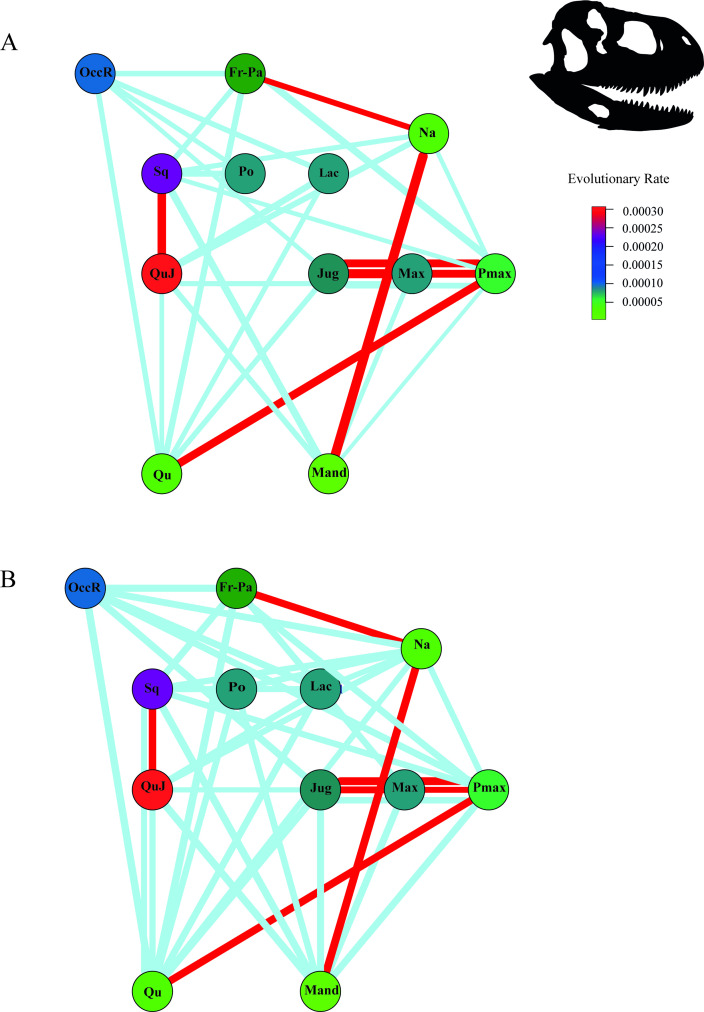
New approaches on the biogeography and phylogeny of abelisaurid theropods
Novas, F.E. & Bandyopadhyay, S. · VII International Symposium on Mesozoic Terrestrial Ecosystems, Buenos Aires
Trabalho de conferência que antecede a descrição formal do Rajasaurus e estabelece o quadro biogeográfico e filogenético para os abelissaurídeos gondwânicos, incluindo material indiano que séria posteriormente formalizado. Novas e Bandyopadhyay discutem as conexões evolutivas entre os abelissaurídeos da Índia e de Madagascar, propondo que a separação final desses dois landmasses ocorreu após o estabelecimento de uma fauna abelissaurídea compartilhada. Este trabalho preparou conceitualmente o campo para a publicação formal de Wilson et al. (2003) e estabeleceu o enquadramento biogeográfico dentro do qual o Rajasaurus é interpretado como representante do isolamento evolutivo da placa indiana durante o Cretáceo Superior.


New theropod remains from Lameta Formation (Maastrichtian) of Jabalpur, India
Srivastava, S., Bhatt, D.K. & Khosla, A. · Current Science
Relatório preliminar sobre novos restos de terópodes da Formação Lameta de Jabalpur, representando parte do material que séria subsequentemente descrito formalmente como Rajasaurus narmadensis em 2003. Srivastava, Bhatt e Khosla descrevem vértebras e elementos do membro posterior que formam o núcleo da futura coleção do holótipo. Este trabalho representa a primeira documentação sistemática do material que levaria à descoberta formal do Rajasaurus, e é fundamental para entender a história de coleta do espécime entre as campanhas de escavação de 1982-1984 e a descrição formal de 2003. Os autores reconhecem a afinidade abelissaurídea do material, mas sem suficiente material craniano para uma descrição formal completa naquele momento.


Rajasaurus narmadensis: a new Indian dinosaur
Wilson, J.A. · Current Science
Comunicação científica sumária anunciando a descrição formal do Rajasaurus narmadensis para a comunidade científica indiana, publicada na Current Science — a principal revista científica de amplo espectro da Índia. Wilson resume os caracteres diagnósticos do novo abelissaurídeo e contextualiza sua posição filogenética entre os terópodes gondwânicos. Este artigo teve papel fundamental na divulgação da descoberta para a comunidade científica indiana e para o público geral, contribuindo para o reconhecimento do Rajasaurus como uma das espécies mais importantes da paleontologia indiana. A visibilidade gerada por esta comunicação foi central para o subsequente estabelecimento de um parque de dinossauros em Rahioli e a erração de uma estátua do animal na localidade de descoberta.


The 'face' of the Triassic: an exceptional case of convergence in basal iguanodontians
Sales, M.A.F., Lacerda, M.B., Horn, B.L.D., de Oliveira, I.A.P. & Schultz, C.L. · PLOS ONE
Análise da distribuição global de três famílias de terópodes — Abelisauridae, Carcharodontosauridae e Spinosauridae — ao longo de três períodos de tempo, mapeando a distribuição paleogeográfica dessas famílias em mapas paleocontinentais do Jurássico Superior, Cretáceo Inferior e Cretáceo Superior. A ocorrência do Rajasaurus narmadensis na Índia representa um ponto de dados chave para entender a dispersão dos abelissaurídeos a partir de Gondwana. O estudo demonstra que os abelissaurídeos mantiveram uma distribuição centrada em Gondwana ao longo do Cretáceo, com o isolamento da placa indiana durante o Cretáceo Superior resultando em uma linhagem endêmica que culminou no Rajasaurus.




Espécimes famosos em museus
GSI 21141/1-33 (Holótipo)
Geological Survey of India, Jaipur / Regional Museum of Natural History, Bhopal
O holótipo GSI 21141/1-33, escavado em Rahioli, Gujarat entre 1982 e 1984, é o único espécime formal do Rajasaurus narmadensis e representa o primeiro terópode indiano com restos pós-cranianos significativos. Inclui partes do crânio, vértebras, quadril e membros posteriores.
Réplica exposta (Regional Museum of Natural History)
Regional Museum of Natural History, Bhopal, Madhya Pradesh, Índia
Uma réplica completa do Rajasaurus está em exibição permanente no Museu Regional de História Natural de Bhopal, Madhya Pradesh. O museu expõe tanto a réplica do esqueleto quanto modelos de vida, tornando-se um dos principais centros de divulgação da paleontologia do Cretáceo indiano para o público.
No cinema e na cultura popular
O Rajasaurus narmadensis teve uma trajetória de representação cultural marcada pelo anonimato inicial e pelo reconhecimento tardio. Por duas décadas após sua descrição formal em 2003, o dinossauro permaneceu quase invisível na cultura popular global, apesar de ser o maior predador já descoberto no subcontinente indiano. Sua estreia televisiva mais significativa veio somente em 2023, no episódio 'Badlands' de Prehistoric Planet (Apple TV+), onde foi retratado com uma coloração dramática de vermelho e preto adaptada ao ambiente vulcânico das Armadilhas do Decã — uma escolha artística ousada que rapidamente se tornou a imagem mais reconhecida da espécie. Na Índia, o Rajasaurus tem um papel cultural diferente: é símbolo do orgulho paleontológico nacional, com uma atração dedicada no parque Adlabs Imagica e uma estátua na localidade de descoberta em Rahioli, Gujarat. O game Jurassic World Evolution 2 e o mobile Jurassic World Alive popularizaram o nome para audiências de jogos globalmente. A conexão biogeográfica com o Majungasaurus de Madagascar, amplamente documentada, tornou o Rajasaurus um personagem recorrente nas discussões sobre deriva continental e evolução convergente em documentários científicos.
Classificação

Descoberta
Curiosidade
O Rajasaurus foi o único abelissaurídeo da Índia com crista craniana mediana — uma protuberância óssea única sobre o focinho, formada pelos ossos nasofrontais. Quando a Índia era uma ilha à deriva no oceano em direção à Ásia, esse predador singular evoluiu isolado por milhões de anos, tornando-se parente distante do Majungasaurus de Madagascar e do Carnotaurus da Argentina — todos primos gondwânicos separados pela deriva continental.
Última revisão: 24 de abril de 2026