Brontossauro
Brontosaurus excelsus
"Lagarto do trovão nobre"
Sobre esta espécie
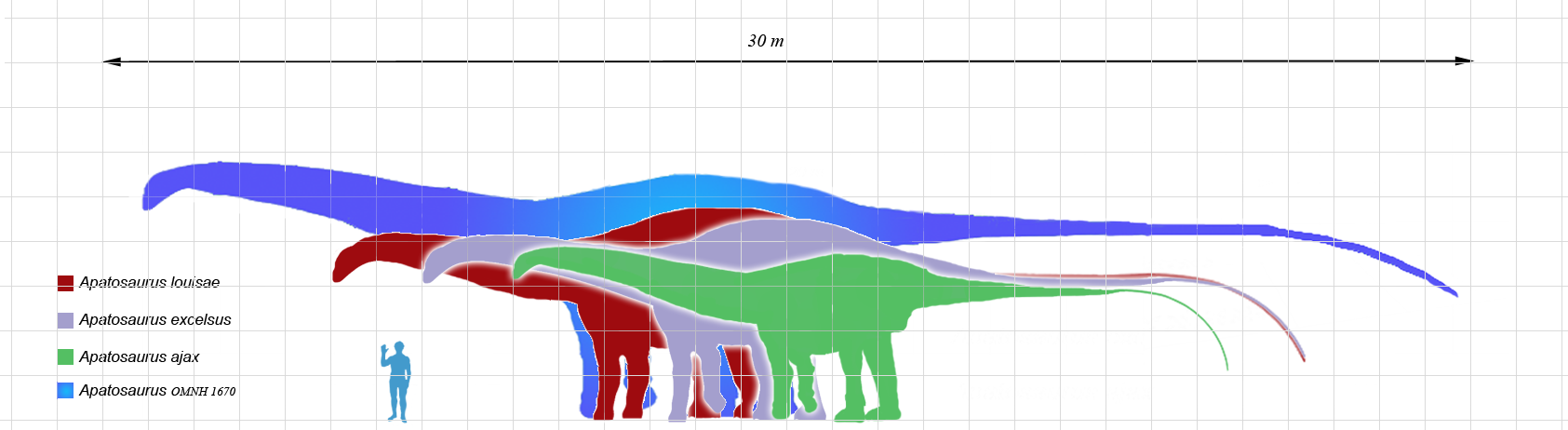
O Brontosaurus excelsus é um dos saurópodes mais icônicos da história da paleontologia. Viveu no Jurássico Superior, há 156 a 146 milhões de anos, nas planícies semiaridas do que hoje é o oeste dos Estados Unidos. Com até 22 metros de comprimento e aproximadamente 17 toneladas, era um dos maiores herbívoros de seu ecossistema. Seu pescoço longo e robusto, muito mais espesso que o de seus parentes diplodocídeos como Diplodocus, pode ter servido para combate intraespecífico além de alimentação. Sinonimizado com Apatosaurus em 1903 por Elmer Riggs, o gênero foi revalidado em 2015 por Tschopp, Mateus e Benson após a maior análise filogenética já realizada sobre diplodocídeos.
Formação geológica e ambiente
A Formação Morrison é uma das sequências sedimentares mais importantes para a paleontologia de dinossauros no mundo. Depositada entre 156 e 147 milhões de anos atrás no Jurássico Superior (idades Kimmeridgiana a Titoniana), cobre grande parte do oeste dos Estados Unidos, do Novo México ao Montana. O ambiente era uma ampla planície aluvial semiarida com estações secas e úmidas pronunciadas, cruzada por rios sazonais e pontilhada de lagos rasos. A formação preservou a maior diversidade de sauropodes já conhecida em um único ecossistema: mais de dez gêneros, incluindo Diplodocus, Brachiosaurus, Camarasaurus, Apatosaurus e o próprio Brontosaurus excelsus.
Galeria de imagens











Reconstituição científica de Brontosaurus excelsus por Nobu Tamura (2016), mostrando o animal em postura lateral com pescoço robusto e cauda levantada, coerente com o consenso científico pós-2015.
Nobu Tamura — CC BY-SA 4.0
Ecologia e comportamento
Habitat
Brontosaurus excelsus habitava as planícies aluviais e florestas de coníferas da Formação Morrison, um ambiente semiarido com estações úmidas e secas distintas no que hoje é o oeste dos Estados Unidos. O ecossistema era dominado por samambaias, cicas, ginkgos, cavalinhas e coníferas como Brachyphyllum, sem gramíneas florescentes. Rios sazonais, lagoas rasas e planícies de inundação forneciam água e concentravam a vegetação mais densa. Compartilhava o habitat com outros sauropodes gigantes como Diplodocus, Apatosaurus, Brachiosaurus e Camarasaurus, além de terópodes como Allosaurus e Ceratosaurus e ornitísquios como Stegosaurus.
Alimentação
Herbívoro estrito, Brontosaurus excelsus alimentava-se de vegetação de baixa a média altura utilizando seus dentes simples em forma de lápis, concentrados na parte frontal de um focinho largo — adaptação para pastejo não seletivo em ampla área. Análises de microwear dentário em diplodocídeos sugerem que o animal varreria a vegetação com movimentos laterais da cabeça, consumindo grandes volumes de samambaia, cavalinhas e folhagem baixa de coníferas. O pescoço excepcionalmente robusto do Brontosaurus, distinto do pescoço mais grácil de Diplodocus, pode ter permitido acesso a vegetação mais resistente ou densa. Estimativas de consumo sugerem que um adulto precisava de centenas de quilogramas de vegetação por dia.
Comportamento e sentidos
As evidências de comportamento de Brontosaurus excelsus são inferidas por comparação com parentes e por morfologia funcional. O pescoço excepcionalmente robusto, com costelas cervicais ventralmente deslocadas e ramos parapofiseais ventrolateralmente orientados, foi interpretado por Taylor et al. (2015) como possível adaptação para combate pescoço a pescoço entre machos, analogamente ao comportamento de 'necking' das girafas. Se confirmada, essa hipótese implicaria comportamento social complexo com competição intraespecífica por acesso a fêmeas. Não há evidência direta de comportamento de rebanho, mas a abundância de espécimes em algumas localidades sugere que indivíduos ocupavam áreas de alimentação comuns.
Fisiologia e crescimento
Brontosaurus excelsus apresentava o esqueleto axial altamente pneumatizado típico dos sauropodes: vértebras com cavidades internas preenchidas por divertículos de sacos aéreos semelhantes ao sistema aviário, reduzindo o peso do pescoço e tronco sem sacrificar a resistência estrutural. King et al. (2024) descreveram recentemente pneumaticidade também nas costelas dorsais — extensão inédita do sistema de sacos aéreos nos apatossaurinos. A histologia óssea indica crescimento rápido e sustentado na juventude, sugerindo metabolismo elevado. Estudos de Curry (1999) e Lehman & Woodward (2008) estimam que o animal atingia tamanho adulto em 15 a 70 anos, dependendo do modelo. A termorregulação provavelmente se beneficiava da gigantotermia: o volume corporal enorme retinha calor metabólico mesmo sem endotermia plena.
Paleogeografia
Configuração continental

Ron Blakey · CC BY 3.0 · Jurássico, ~90 Ma
Durante o Kimmeridgiano–Titoniano (~156–146 Ma), Brontosaurus excelsus habitava a Pangeia em processo de fragmentação. A América do Norte e a Europa ainda estavam próximas, e o Atlântico Norte mal começava a se abrir. O clima era quente e úmido em escala global, sem calotas polares.
Inventário de Ossos
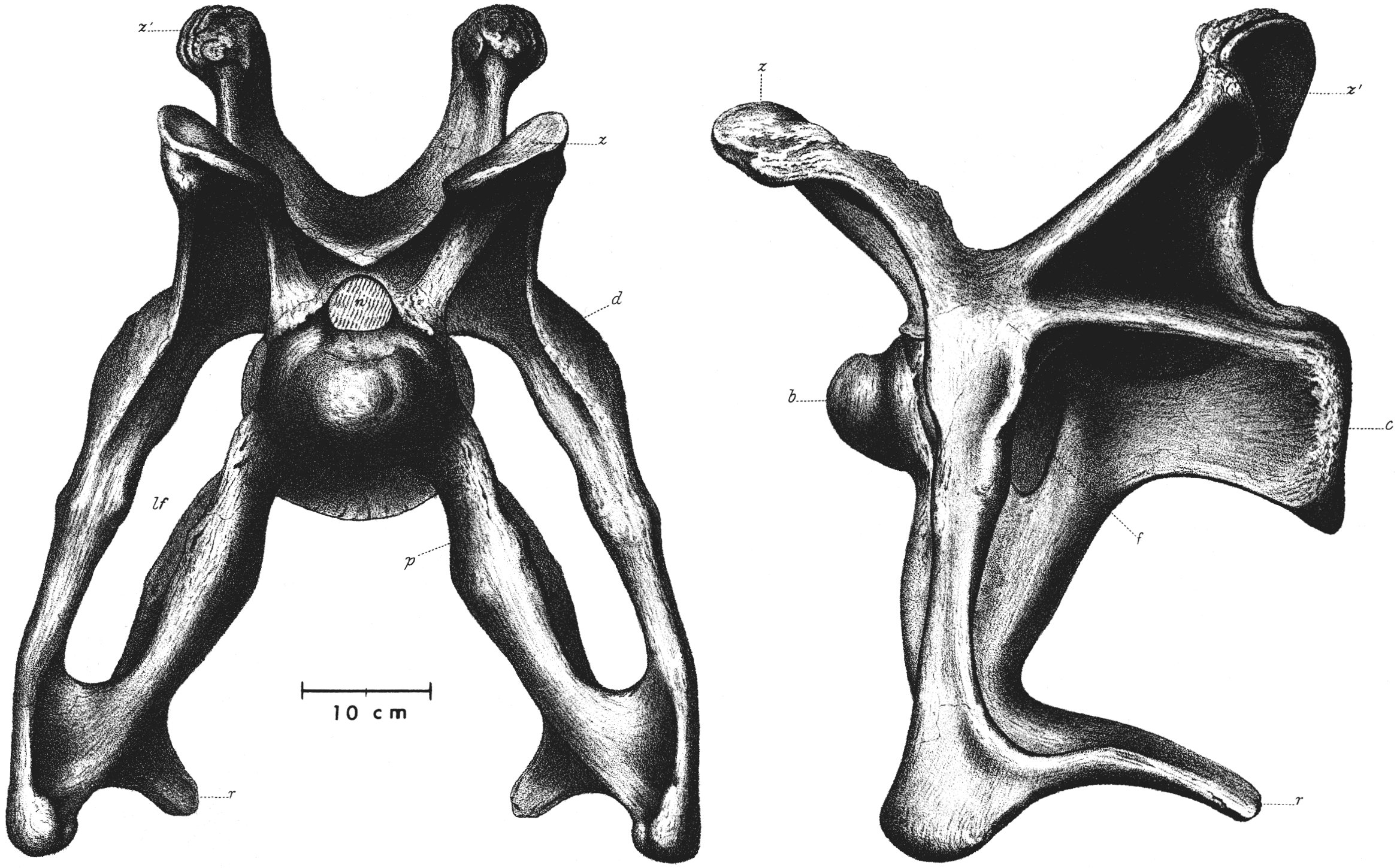
O holótipo YPM 1980, coletado em Como Bluff (Wyoming) em 1879, preserva vértebras, costelas, cintura peitoral e membros, mas carecia de crânio. O espécime AMNH 460 (montado em 1905 no Museu Americano de História Natural) é um composto de vários indivíduos. O crânio verdadeiro do grupo só foi estabelecido em 1975 por McIntosh e Berman, que demonstraram ser semelhante ao de Diplodocus.
Estruturas encontradas
Estruturas inferidas
Literatura Científica
15 artigos em ordem cronológica — do artigo de descrição original até pesquisas recentes.
Notice of New Jurassic Reptiles
Marsh, O.C. · American Journal of Science
Artigo fundador em que Othniel Charles Marsh nomeia Brontosaurus excelsus com base no espécime YPM 1980, coletado por William Harlow Reed em Como Bluff, Wyoming. A descrição, publicada no American Journal of Science em dezembro de 1879, estabelece o gênero com base em vértebras cervicais e dorsais, costelas, cintura peitoral e membros posteriores. Marsh distingue Brontosaurus de Apatosaurus (descrito dois anos antes) por proporções diferentes das vértebras e membros, e aponta o tamanho colossal como excepcional. O nome Brontosaurus — 'lagarto do trovão', do grego brontē e sauros — e o epíteto excelsus, do latim 'nobre' ou 'elevado', refletem a grandiosidade percebida do animal. Este artigo inaugura uma das maiores rivalidades científicas da história: a Bone Wars entre Marsh e Edward Drinker Cope, e lança um dos nomes mais famosos da paleontologia.
The Dinosaurs of North America
Marsh, O.C. · 16th Annual Report of the U.S. Geological Survey
Obra monumental de Marsh publicada como relatório anual do U.S. Geological Survey, apresentando a primeira reconstituição esquelética completa de Brontosaurus excelsus. A ilustração, baseada no holótipo YPM 1980 complementado por material de outros espécimes da Morrison Formation, mostra o animal em postura ereta com cauda levemente erguida, definindo a imagem canônica do saurópode para o grande público e para a ciência da época. Marsh inclui comparações com outros dinossauros norte-americanos, discute as relações filogenéticas dos diplodocídeos e propõe interpretações paleoecológicas do ecossistema da Formação Morrison. O trabalho se torna referência obrigatória para todo estudo subsequente sobre sauropomorfos jurássicos da América do Norte, e a reconstituição esquelética é reproduzida em livros didáticos e museus ao redor do mundo por décadas.
Structure and Relationships of Opisthocoelian Dinosaurs. Part I, Apatosaurus Marsh
Riggs, E.S. · Publications of the Field Columbian Museum, Geological Series
Paper decisivo em que Elmer Riggs, do Field Museum de Chicago, compara o espécime de Brontosaurus excelsus de Como Bluff com o novo esqueleto P25112 (Apatosaurus) coletado em Fruita, Colorado. Riggs conclui que as características diagnosticadas por Marsh para separar Brontosaurus de Apatosaurus — proporções vertebrais e de membros — na verdade refletem diferenças ontogenéticas, sendo o holótipo de Apatosaurus ajax um indivíduo imaturo. Assim, pelo princípio de prioridade nomenclatural, Apatosaurus (1877) prevalece sobre Brontosaurus (1879), e Riggs cria a combinação Apatosaurus excelsus. Publicado em um periódico relativamente obscuro, o argumento de Riggs levou décadas para ser plenamente assimilado pela comunidade científica e pelo público. A sinonímia iniciaria 112 anos de controvérsia que só seria resolvida pelo trabalho de Tschopp et al. em 2015.
Description of the Palate and Lower Jaw of the Sauropod Dinosaur Diplodocus (Reptilia: Saurischia) with Remarks on the Nature of the Skull of Apatosaurus
McIntosh, J.S. & Berman, D.S. · Journal of Paleontology
Estudo revolucionário que corrige um erro de 70 anos na reconstituição do Brontosaurus/Apatosaurus. Desde 1905, os montes museológicos exibiam crânios do tipo Camarasaurus — robusto, com dentes espatulados — porque nenhum crânio havia sido encontrado com os esqueletos pós-cranianos. McIntosh e Berman analisam os espécimes de crânio disponíveis e demonstram que a morfologia craniana correta é a diplodocídea: crânio estreito, baixo, com dentes simples em forma de lápis concentrados na frente da boca. A descoberta de um crânio associado ao espécime CM 3018 (Carnegie Museum) em 1909, que permanecera sem estudar por décadas, confirma a conclusão. A rectificação é adotada pelo Carnegie Museum em 1979, quando o primeiro crânio correto é montado em um esqueleto de apatossaurino — quatro anos depois da publicação deste paper.
Supersonic Sauropods? Tail Dynamics in the Diplodocids
Myhrvold, N.P. & Currie, P.J. · Paleobiology
Estudo pioneiro de Nathan Myhrvold e Philip Currie que usa simulação computacional para modelar a dinâmica da cauda de Apatosaurus louisae — táxon intimamente relacionado ao Brontosaurus excelsus. Os resultados sugerem que a cauda longa e afilada dos diplodocídeos podia ser 'estralada' como um chicote, com a ponta atingindo velocidades supersônicas e produzindo um estouro sonoro análogo ao de um chicote real. A hipótese propõe que esse comportamento servia a funções comunicativas ou defensivas. Publicado em Paleobiology em 1997, o artigo gerou enorme repercussão científica e midiática, embora análises posteriores (Parrish et al., 2013; Gutiérrez-Blanco et al., 2022) tenham contestado se as velocidades supersônicas eram realmente atingíveis dado o atrito muscular e a resistência do ar. Mesmo refutado parcialmente, o paper estabeleceu um novo paradigma na paleoetologia dos saurópodes.
Ontogenetic histology of Apatosaurus (Dinosauria: Sauropoda): new insights on growth rates and longevity
Curry, K.A. · Journal of Vertebrate Paleontology
Estudo histológico clássico de Kristina Curry que analisa a microestrutura óssea de uma série ontogenética de rádios, ulnas e escapulas de Apatosaurus — gênero diretamente relacionado ao Brontosaurus excelsus dentro de Apatosaurinae. O exame de linhas de crescimento (LAGs) e tecido ósseo fibrolamelar revela três fases osteogênicas distintas: crescimento rápido e sustentado durante a maior parte da ontogenia, desaceleração gradual ao aproximar-se do tamanho adulto, e eventual cessação. A estimativa de ~10 anos para sub-adultos grandes refuta a hipótese de que crescimento lento e indeterminado seria necessário para os sauropodes atingirem tamanhos extremos. O trabalho inaugurou a aplicação sistemática de histologia óssea a sauropodes e estabeleceu o paradigma de crescimento rápido 'à moda das aves' para o grupo.
Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs
Stevens, K.A. & Parrish, J.M. · Science
Estudo seminal de Kent Stevens e Michael Parrish que usa reconstituições digitais articuladas dos pescoços de Apatosaurus e Diplodocus — táxons fundamentais para compreender a ecologia de Brontosaurus excelsus — para inferir a postura cervical neutra. Utilizando o programa DinoMorph, os autores modelam as articulações intervertebrais e concluem que ambos os diplodocídeos mantinham o pescoço em posição levemente inclinada para baixo em repouso, com a cabeça próxima ao nível do solo. O resultado sugere alimentação a baixa altura — gramíneas, samambaias e vegetação rasteira — e não pastejo de copas de árvores. O paper gerou debate intenso: Taylor et al. (2009) argumentariam que a postura neutra em animais terrestres vivos é a elevação máxima sustentável, implicando pescoços erguidos. A controvérsia moldou duas décadas de pesquisa em paleoecologia de sauropomorfos.

Modeling growth rates for sauropod dinosaurs
Lehman, T.M. & Woodward, H.N. · Paleobiology
Lehman e Woodward analisam seções histológicas de ossos longos de Apatosaurus — diretamente relacionado ao Brontosaurus excelsus — combinando contagem de linhas de crescimento (LAGs) com estimativas de massa corporal para modelar curvas de crescimento. Dois modelos alternativos são testados: um baseado na relação entre comprimento ósseo e massa corporal que sugere atingimento do tamanho adulto (~25.000 kg) em apenas 15 anos com taxa máxima de crescimento de 5.000 kg/ano; e o modelo de von Bertalanffy que projeta 70 anos até o tamanho adulto com taxa máxima de 520 kg/ano. A discrepância reflete incertezas fundamentais sobre a relação entre histologia, massa e metabolismo em gigantes jurássicos. O paper é referência central no debate sobre se sauropodes eram endotérmicos, ectotérmicos ou mesotérmicos.

Why sauropods had long necks; and why giraffes have short necks
Taylor, M.P. & Wedel, M.J. · PeerJ
Taylor e Wedel analisam por que os pescoços dos sauropodes — incluindo Brontosaurus excelsus, que possuía um pescoço notavelmente robusto para um diplodocídeo — atingiram comprimentos de até 15 metros, superando em seis vezes o pescoço mais longo de qualquer girafa. O paper avalia sistematicamente seis hipóteses: acesso a vegetação de alta copa, alimentação em superfícies aquáticas, varrimento de área ampla sem mover o corpo, termorregulação, seleção sexual e evolução neutra. A conclusão é que a vantagem alimentar — acesso exclusivo a recursos vegetais fora do alcance de outros herbívoros — é o principal fator seletivo, com seleção sexual como fator secundário plausível. A análise enfatiza que os pescoços de Apatosaurus/Brontosaurus são morfologicamente distintos dos de Diplodocus e Brachiosaurus, sugerindo estratégias de uso diferentes dentro do mesmo ecossistema.


Inferences of Diplodocoid (Sauropoda: Dinosauria) Feeding Behavior from Snout Shape and Microwear Analyses
Whitlock, J.A. · PLOS ONE
Whitlock combina análise de forma do focinho e microscopia de desgaste dentário (microwear) para inferir o comportamento alimentar dos diplodocóideos, grupo que inclui Brontosaurus excelsus. O estudo identifica dois estratégias alimentares distintas: diplodocídeos com focinhos quadrados realizavam pastejo não seletivo em ampla área ao nível do solo, enquanto braquiossaurídeos com focinhos arredondados e mais robustos exerciam pastejo seletivo em maior altura. Brontosaurus/Apatosaurus, com seu focinho intermediário e pescoço excepcionalmente robusto, parece ter ocupado nicho ecológico distinto dos demais membros da família. O paper fornece a primeira evidência quantitativa baseada em microwear para partição de nicho alimentar entre os gigantes herbívoros da Formação Morrison.


Were the necks of Apatosaurus and Brontosaurus adapted for combat?
Taylor, M.P., Wedel, M.J., Naish, D. & Engh, B. · PeerJ PrePrints
Taylor, Wedel, Naish e Engh examinam características morfológicas do pescoço de Apatosaurus e Brontosaurus — especificamente o deslocamento ventral das costelas cervicais e a orientação ventrolateral dos ramos parapofiseais — e argumentam que essas estruturas são consistentes com adaptações para combate intraespecífico pescoço a pescoço, analogamente ao 'necking' observado em girafas. O pescoço excepcionalmente robusto e musculoso de Brontosaurus, em contraste com o pescoço mais grácil de Diplodocus, é visto como evidência de que diferentes gêneros de diplodocídeos utilizavam o pescoço para funções diferentes. A hipótese de combate implicaria que Brontosaurus tinha comportamento social mais complexo do que anteriormente suposto, com machos competindo por acesso a fêmeas ou recursos. O paper é um preprint PeerJ e representa uma das hipóteses comportamentais mais discutidas sobre sauropodes.


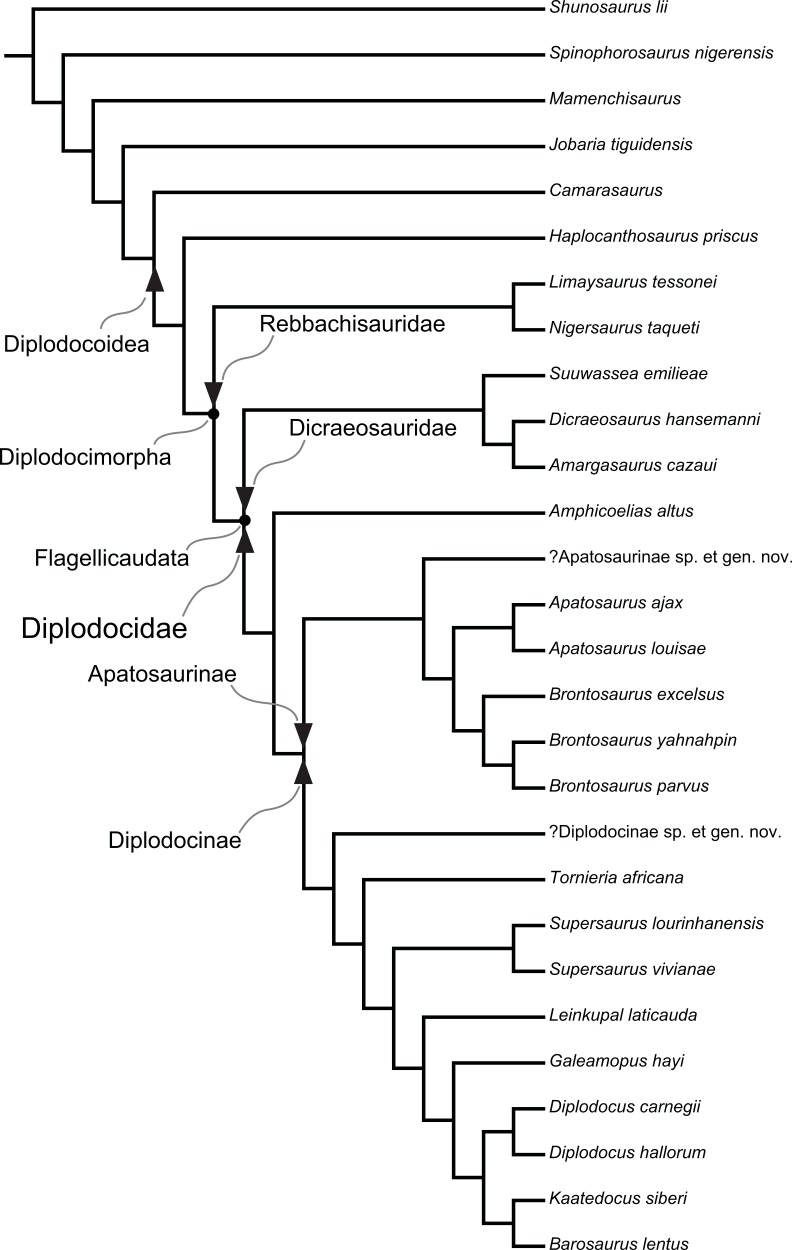
A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)
Tschopp, E., Mateus, O. & Benson, R.B.J. · PeerJ
O paper de revalidação de Brontosaurus, publicado em PeerJ em 2015, é o trabalho filogenético mais abrangente já realizado sobre Diplodocidae. Tschopp, Mateus e Benson pontuam 477 caracteres morfológicos em 81 espécimes de sauropodes — uma análise de cinco anos envolvendo visitas a museus na Europa e nos Estados Unidos. O método inovador de análise em nível de espécime (ao invés do nível de espécie) permite detectar diferenças sutis entre táxons historicamente confundidos. O resultado é inequívoco: Brontosaurus apresenta suficientes autapomorfias (características exclusivas) que o distinguem de Apatosaurus, revalidando o gênero após 112 anos de sinonímia. Três espécies são reconhecidas: B. excelsus (tipo), B. parvus e B. yahnahpin. O paper gerou cobertura midiática global e é o artigo científico de paleontologia de vertebrados mais lido de 2015.


Evidence for niche partitioning among ground-height browsing sauropods from the Upper Jurassic Morrison Formation of North America
McHugh, J.B. · Geology of the Intermountain West
McHugh examina evidências isotópicas e morfométricas de sauropodes da Formação Morrison para investigar como múltiplos gêneros gigantes — incluindo Brontosaurus excelsus — coexistiam no mesmo ecossistema sem competição alimentar direta. O estudo foca em pastadores a baixa altura, propondo que diferenças na morfologia dentária, formato do focinho e amplitude de movimentação do pescoço permitiam a cada gênero explorar microhabitats alimentares distintos. Brontosaurus, com seu pescoço excepcionalmente robusto e musculoso, pode ter explorado vegetação densa ou resistente inacessível aos demais diplodocídeos. O artigo de acesso aberto contribui para resolver o 'paradoxo da diversidade da Morrison' — a extraordinária coexistência de mais de dez gêneros de sauropodes em um ambiente semiarido com recursos alimentares limitados.


A previously unreported form of dorsal rib pneumaticity in Apatosaurus and Brontosaurus
King, A., Wedel, M.J. & Taylor, M.P. · PeerJ
King, Wedel e Taylor descrevem, pela primeira vez, pneumaticidade em costelas dorsais de Apatosaurus e Brontosaurus — um tipo de cavidade aérea interna semelhante às encontradas em aves, estendida para além das vértebras (já conhecidas como pneumáticas) até as costelas. O estudo usa tomografia computadorizada e exame de superfície de espécimes em coleções museológicas para documentar câmaras e fossas pneumáticas dentro do tecido ósseo costal. A descoberta implica que o sistema de sacos aéreos — fundamental para ventilação pulmonar eficiente em aves — era ainda mais extenso nos apatossaurinos do que se pensava, com divertículos penetrando o esqueleto axial de forma ainda mais abrangente. Este é um dos papers mais recentes sobre a anatomia de Brontosaurus, publicado em 2024 no PeerJ.


Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs
Wedel, M.J. · Paleobiology
Wedel analisa a pneumaticidade vertebral dos sauropodes — um sistema de câmaras e fossas internas nas vértebras preenchidas por divertículos de sacos aéreos semelhante ao sistema das aves modernas. O estudo documenta a distribuição filogenética da pneumaticidade em sauropodes: basais apenas nas vértebras pré-sacrais, derivados (como diplodocídeos, incluindo Brontosaurus excelsus) com extensão até as caudais anteriores. A presença de sacos aéreos análogos aos das aves implica um sistema respiratório de fluxo contínuo de ar — muito mais eficiente que o pulmão bidirecional dos mamíferos — e é consistente com altas taxas metabólicas. O paper estabeleceu a pneumaticidade vertebral como ferramenta diagnóstica e fisiológica fundamental na paleobiologia de sauropodes, e é o trabalho de referência que antecede a descoberta de King et al. (2024) sobre a pneumaticidade nas costelas de Brontosaurus.


Espécimes famosos em museus
YPM 1980 (Holótipo)
Yale Peabody Museum of Natural History, New Haven, Connecticut, EUA
Holótipo da espécie, coletado em Como Bluff, Wyoming. Preserva vértebras cervicais e dorsais, costelas, cintura peitoral, membros anteriores e posteriores, mas não inclui crânio. Está em exibição permanente no Great Hall do Peabody Museum desde 1931, com 20 metros de comprimento montado.
AMNH 460 (Monte Composto)
American Museum of Natural History, Nova York, EUA
Monte histórico inaugurado em 1905 como o primeiro esqueleto de sauropode do mundo. Composto principalmente do espécime AMNH 460 do Nine Mile Crossing Quarry (Como Bluff), suplementado por ossos de AMNH 592, 222 e 339 e moldes em gesso do holótipo YPM 1980. Tschopp et al. (2015) classificaram este espécime como 'apatossaurino indeterminado' pendente análise detalhada.
No cinema e na cultura popular
Brontosaurus é, talvez, o dinossauro mais presente no imaginário cultural do século XX. O nome entrou para o vocabulário popular antes mesmo de ter sido sinonimizado com Apatosaurus em 1903 — e permaneceu como sinônimo de 'dinossauro de pescoço longo' na mente do público por mais de um século, independentemente de qualquer revisão taxonômica. No cinema, a trajetória começa em 1925 com 'The Lost World', onde um Brontosaurus é o personagem central do clímax londrino, e continua em 1933 com 'King Kong', onde o animal aparece como predador temível em Skull Island. Em 1940, Disney o imortalizou em 'Fantasia' pastejando em paisagens jurássicas ao som de Stravinsky. Décadas depois, em 1993, 'Jurassic Park' menciona o nome como uma confusão de criança — reflexo preciso de como Brontosaurus vivia no vocabulário popular mesmo quando era cientificamente sinônimo de Apatosaurus. Em 2015, o filme de animação 'O Bom Dinossauro' da Pixar protagonizou um apatossaurino no mesmo ano em que Tschopp et al. revalidavam o gênero. A revalidação científica de 2015 fechou um ciclo de 112 anos: Brontosaurus voltou a ser um nome válido — e a cultura pop, que nunca o abandonou, pode finalmente dizer que sempre teve razão.

Classificação

Descoberta
Curiosidade
Brontosaurus foi considerado extinto duas vezes: primeiro como animal real (há 146 milhões de anos) e depois como nome científico válido — suprimido por 112 anos (1903–2015) até ser 'ressuscitado' pelo maior estudo filogenético já realizado sobre diplodocídeos. É o único dinossauro da história a ter seu gênero oficialmente abolido e depois restaurado.